Frontiers in Systems Neuroscience
Änderungen des Hirnzustands sind mit Veränderungen der Mikroschaltkreise in den neuronalen Feuereigenschaften und mit Veränderungen auf der Makroebene in synchronen oder asynchronen Mustern der Hirnaktivierung verbunden. In diesem Rahmen wurden Axone relativ wenig untersucht (Barry, 2015), und zwar hauptsächlich im Zusammenhang mit der Leitungsgeschwindigkeit von Aktionspotenzialen und der erhöhten Wachsamkeit, die sich aus Änderungen der Leitungsgeschwindigkeit ergeben kann (z. B. Stoelzel et al., 2017). Darüber hinaus scheint die stark verzweigte Topologie vieler Axone jedoch gut dafür geeignet zu sein, verteilte postsynaptische Neuronengruppen differenziert zu rekrutieren, möglicherweise in einer zustandsabhängigen Weise. Die Signalverarbeitung kann in mindestens drei axonalen Bereichen erfolgen, nämlich bei der Zuordnung, der Verstärkung und dem Timing (Innocenti et al., 2016; Innocenti, 2017). In dieser Perspektive diskutiere ich zunächst kurz Netzwerkbeispiele für Axon-Kollateralisierung und dann einige der axoninternen Merkmale, die der unterschiedlichen postsynaptischen Rekrutierung zugrunde liegen könnten. In Ermangelung detaillierter Parameter oder Mechanismen geht es vor allem darum, allgemeine Merkmale hervorzuheben, die bei der Steuerung und den Übergängen von Gehirnzuständen eine Rolle spielen könnten.
- Axonenkollaterale
- Nicht-stereotypische Kollateralisierung
- Intrinsische Kollaterale
- Ein interessantes Beispiel: Meynert-Zellen im Areal V1 von nicht-menschlichen Primaten
- Funktionelle Bedeutung der Axonverzweigung
- Dynamische Axoneigenschaften
- Schlussfolgerung
- Autorenbeiträge
- Finanzierung
- Conflict of Interest Statement
Axonenkollaterale
Alle Axone haben eine ausgeprägte distale Arborisierung in der Zielstruktur. Viele Axone haben darüber hinaus mehrere Verzweigungen (auch Kollateralen genannt), die auf deutlich unterschiedliche Strukturen abzielen (besprochen in Rockland, 2013, 2018). Ein Paradebeispiel ist die Verzweigung von Axonen aus kortikothalamischen Neuronen der Schicht 5. Dies wurde wiederholt dokumentiert und wird häufig als ein Schlüsselmechanismus diskutiert, durch den ein Organismus unterscheiden kann, ob Veränderungen des sensorischen Inputs durch Veränderungen in der Umgebung oder durch selbst initiierte Bewegungen hervorgerufen werden („Efferenzkopie“ oder „kollaterale Entladung“, Guillery und Sherman, 2011).
Bei Nagetieren, für die es eine Vielzahl von anwendbaren Techniken zur Untersuchung dieses Problems gibt, ist die Kollateralisierung bekanntlich relativ häufig. Um einige Beispiele zu nennen: (1) Neuronen im hippocampalen Subiculum projizieren zu den Mammillarkörpern, dem retrosplenialen Kortex oder über Kollateralen zu beiden (Ratte: Kinnavane et al, 2018); (2) serotonerge Neuronen im dorsalen Raphe-Kern (DR-Kern) stehen in verschiedenen Kombinationen mit dem Striatum, dem präfrontalen Kortex und der Amygdala in Verbindung (Ratte: Gagnon und Parent, 2014) sowie mit Kombinationen von autonomen Kernen (Waselus et al., 2011). In diesem Fall könnte die Aktivierung der Stressreaktion durch DR-Kollaterale zu einer synchronisierten Aktivierung von Kernen führen, die mit der Freisetzung von Neurohormonen oder Druckreaktionen in Verbindung stehen. Unterschiedliche, koordinierte Aktivierungen von Vorderhirnstrukturen könnten zu den vielfältigen, aber verwandten DR-Funktionen wie der Regulierung des Schlaf-Wach-Zyklus, der Modulation von Schmerzsignalen oder der Stimmungsäußerung beitragen (Gagnon und Parent, 2014).
Drittens wurden in anatomischen Studien der thalamokortikalen Projektionen von Mäusen „multispezifische Axone“ identifiziert, die sich weit zu begrenzten Bereichen in separaten kortikalen (und subkortikalen) Bereichen verzweigen. Es wurde angenommen, dass diese die schnelle Entstehung und Rekonfiguration räumlich verteilter, synchronisierbarer neuronaler Verbände orchestrieren (Clasca et al., 2016).
Viertens ergab eine kürzlich durchgeführte Studie über kortiko-kortikale Verbindungen unter Verwendung der axonalen Verfolgung des gesamten Gehirns im visuellen Kortex der Maus, dass 23 von 30 Neuronen mit zwei bis sieben anderen kortikalen Bereichen in Kontakt standen. Zur Bestätigung dieses Ergebnisses ergab die Hochdurchsatz-DNA-Sequenzierung von genetisch barcodierten Neuronen, dass 44 % von 533 Neuronen mehrfach projizieren („broadcast neurons“, Han et al., 2018). Han et al. unterschieden vorläufig zwei Arten von projizierenden Neuronen, eine kleinere „dedizierte“ (uni-target) Subpopulation, die mit einer Prävalenz von „broadcasting“ (mehrfach projizierenden) Zellen koexistiert. Könnte diese Architektur für Modulationen des kognitiven Zustands und der sensorischen Verarbeitung verantwortlich sein?
Nicht-stereotypische Kollateralisierung
Wie bereits erwähnt, erfolgt die Kollateralisierung von Neuronen nach einem nicht-stereotypischen Muster. Innerhalb einer bestimmten Projektion (definiert durch den Ursprung) senden Neuronen Verzweigungen zu einer diversifizierten Teilmenge von Zielgebieten („in allen Kombinationen“). Diese Beobachtung ist so konsistent, dass sie als Regel angesehen werden kann, auch wenn die detaillierten Parameter nicht tabellarisch erfasst wurden. Die funktionelle Bedeutung ist ebenfalls unklar, aber neue Ergebnisse deuten darauf hin, dass die heterogenen und variablen Eingänge zu einem kortikalen Neuron (und im weiteren Sinne auch zu neuronalen Verbänden) wichtig sind, um die Variabilität und die Veränderungen der Spike-Trains über experimentelle Versuche hinweg zu steuern (Gomez-Laberge et al., 2016 und siehe unten).
Intrinsische Kollaterale
In allen Spezies haben kortikale Fernprojektionsneuronen, zusätzlich zu einzelnen oder mehreren extrinsischen Zielen, typischerweise eine ausgeklügelte intrinsische Arborisierung. Einige kortikale Neuronen haben nur eine intrinsische (lokale) Kollateralisation, und einige nur eine extrinsische (Ratte: Kita und Kita, 2012). Obwohl es nur wenige Daten über die tatsächlichen Anteile von intrinsischen und extrinsischen Verzweigungen gibt (Parent et al., 2000; Rockland, 2018), gibt es Hinweise darauf, dass dies sehr variabel sein wird. Selbst innerhalb des Systems der intrinsischen Verbindungen gibt es nicht nur Unterschiede in der Anzahl der Kollateralen und der Anzahl der synaptischen Boutons, sondern ein einzelnes Neuron kann eine Mischung aus myelinisierten und nicht myelinisierten Kollateralen aufweisen (visueller Kortex der Katze: Martin et al., 2014; Koestinger et al., 2017). Eine verzweigungsspezifische Myelinisierung könnte zu einer erhöhten, verzweigungsspezifischen Leitungsgeschwindigkeit führen, obwohl Koestinger et al. vermuten, dass dies eher mit Faktoren wie einer erhöhten Übertragungssicherheit zu tun hat, die vermutlich wiederum verzweigungsspezifisch ist.
Eine kuriose Beobachtung im Zusammenhang mit der Myelinisierung betrifft die Stria von Gennari, das myelinisierte Axonband in Schicht 4B des Primatenareals V1. Da dieses Band aus intrinsischen Kollateralen besteht, ist die übliche Erklärung, dass die Myelinisierung ein Mittel zur Erhöhung der Leitungsgeschwindigkeit über große Entfernungen ist, nicht unmittelbar anwendbar. Lokale Kollateralen benötigen keine (?) Vergrößerung in Bezug auf entfernte Ziele, und tatsächlich scheint eine erhöhte lokale Leitgeschwindigkeit (durch Myelinisierung?) paradox zu sein, wenn man die Synchronität vorhersagen würde. Könnte es sein, dass die Myelinisierung mit anderen Faktoren zusammenhängt, wie z.B. mit plastizitätsbedingten (oder zustandsbedingten) Veränderungen des Axondurchmessers?
Ein interessantes Beispiel: Meynert-Zellen im Areal V1 von nicht-menschlichen Primaten
Diese großen Neuronen an der Grenze der Schichten 5 und 6 projizieren variabel in das extrastriate Areal MT und/oder andere visuelle Assoziationsgebiete und/oder in den Nucleus pulvinaris und/oder in den Colliculus superior und das Prätektum (Weisenhorn et al., 1995; Rockland und Knutson, 2001). Die intrinsische Kollateralisation ist außerordentlich umfangreich und wird auf der Grundlage von subtotalen Rekonstruktionen mit 8,0 mm vom Soma gemessen und umfasst mindestens 800-1.370 Boutons (Abbildung 1). Über die axonale Verzweigung hinweg scheint es eine deutliche Variabilität im Zweigdurchmesser zu geben; d. h. die extrinsischen Zweige, die zum Bereich MT gerichtet sind, sind groß (Durchmesser ~3,0 μm), aber die intrinsischen Zweige und die Zweige, die zum Pulvinar und zum Colliculus superior projizieren, scheinen kleiner zu sein, wie durch Lichtmikroskopie beurteilt wird (vgl. Abbildung 1B (intrinsisch) und Abbildung 1D (extrinsisch zum Bereich MT). Unterschiedliche Durchmesser deuten vermutlich auf Unterschiede im Grad der Myelinisierung und folglich in der Leitungsgeschwindigkeit hin.
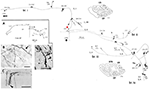
Abbildung 1. Eine typische, räumlich ausgedehnte proximale axonale Verzweigung einer Meynert-Zelle (rotes Sternchen) im primären visuellen Kortex eines Makakenaffen. Es gibt drei intrinsische Hauptkollateralen (beschriftet als br. i, ii, iv), die sich 3,0 mm dorsal in Schicht 1, 3,0 mm ventral in Schicht 6, 4,0 mm ventral in Schicht 6 und 0,5 mm in Schicht 4B erstrecken, wie durch Pfeilspitzen in den beiden koronalen Schnittkonturen (Schnitte 222 und 274, wobei dorsal links ist) angedeutet. Insgesamt erstrecken sich die intrinsischen Kollateralen über 5,9 mm anterior-posterior (117 Schnitte × 50 μm). Eine weitere, extrinsische Kollaterale (br. iii) tritt in den Schichten 4 und 6 des Bereichs V2 auf. Teile der einzelnen Kollateralen und des Hauptaxons (dicker Pfeil) konnten nicht verfolgt werden, was durch gestrichelte Linien angezeigt wird. Der Ausschnitt mit geringer Vergrößerung (A, links) gibt einen schematischen Überblick über die allgemeine Konfiguration. Die Zahlen bezeichnen einzelne Abschnitte, wobei 20 Zahlen = 1,0 mm bedeuten. Alle Äste haben zahlreiche kleine synaptische Cluster, von denen einer in (B) abgebildet ist. Beachten Sie den verringerten Durchmesser zwischen dem Hauptaxon (Pfeil) und der terminalen Verzweigung. Extrinsische Axone (C) sind von unterschiedlichem Durchmesser (ein Axon mit großem Durchmesser in der Pfeilspitze). (D) Extrinsische Endigungen im Bereich MT enthalten einige Axone mit großem Durchmesser. Maßstabsbalken = 25 μm in (B), 100 μm in (C), 20μm in (D). CF, Calcarine-Fissur; LS, Sulcus lunatum; STS, Sulcus temporalis superior; L, Schicht. Modifiziert aus den Abbildungen 1, 9 in Rockland und Knutson (2001) und den Abbildungen 6f, 7b aus Rockland (1995) mit Genehmigung.
Funktionelle Bedeutung der Axonverzweigung
Eine wichtige Komponente der Kollateralisation ist, dass die Tochteräste oft nicht einheitlich sind, sondern insbesondere an den Verzweigungspunkten im Durchmesser variieren (Abbildung 2). Die Variabilität des Durchmessers hat zusammen mit anderen Parametern Auswirkungen auf die Erregbarkeit, die Leitungsgeschwindigkeit und andere Aspekte der Signalausbreitung. Weitere Parameter, die sich auswirken, sind die Breite des Myelins und die intermodale Länge sowie die Dichte und Verteilung der Ionenkanäle (siehe Debanne et al., 2011; Seidl, 2014; Bucher, 2016; Seidl und Rubel, 2016; Rama et al., 2018). Dies hätte Auswirkungen auf die neuronalen Reaktionseigenschaften auf der Ebene der Mikroschaltkreise. Auf der globaleren Ebene könnte die summierte Aktivität mehrerer projizierender Axone mit unterschiedlichen Leitungsgeschwindigkeiten in einem vernetzten Netzwerk zu einem Spektrum synchroner und/oder asynchroner Aktivierungen führen (Mitra et al., 2015; Zeki, 2016). Unterschiedliche Rekrutierung postsynaptischer Populationen oder Netzwerkrekombinationen könnten Faktoren für Zustandsübergänge oder Modulationen sein.
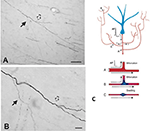
Abbildung 2. Ein typisch verzweigtes Axonsegment in der weißen Substanz (Makakenaffe). Das Segment stammt von einem Neuron im parietalen Kortex und ist hier in der Nähe des ventralen temporalen Kortex zu sehen. Tafel (A) ist eine geringere Vergrößerung von (B). Man beachte die doppelte Verzweigung, wobei der erste Tochterast (durchgezogener Pfeil) auffallend dünner (und unmyelinisiert?) ist als das Hauptaxon. Bei der zweiten, etwas weiter distal gelegenen Verzweigung (hohler Pfeil) scheinen die Tochteräste etwa den gleichen Durchmesser zu haben, aber beide sind dünner als das Hauptaxon. Maßstabsbalken = 100 μm (A) und 10 μm (B). Nachgezeichnet aus Zhong und Rockland (2003) mit Genehmigung. (C) Schematische Darstellung eines Neurons (blau) und seiner erweiterten Verzweigungstopologie (aus Gründen der besseren Formatierung verkürzt). Ein Aktionspotenzial (AP) kann auf verschlungenen Pfaden (A-C) zu mehreren Zielen gelangen. Die Zuverlässigkeit der Ausbreitung hängt sowohl von den aktiven elektrischen Eigenschaften des Axons als auch von seiner Geometrie ab, einschließlich Membraninhomogenitäten wie Schwellungen und inkompatible Astdurchmesser. Unten: Schema zur Veranschaulichung der zuverlässigen Ausbreitung (A, mit optimaler Impedanzanpassung zwischen dem Mutter- und dem Tochterast) und der verlangsamten oder fehlgeschlagenen Ausbreitung (B, wenn der Tochterast einen vergrößerten Durchmesser hat; C, wenn eine Membranschwellung dazwischen liegt). Wiedergegeben aus Huguenard (2000) mit Genehmigung.
Die Axongeometrie, die aktiven elektrischen Eigenschaften und die Membraninhomogenitäten an den Verzweigungspunkten sind als Faktoren für eine zuverlässige Ausbreitung bekannt (z. B. Manor et al., 1991; Innocenti et al., 1994; Tettoni et al., 1998; Huguenard, 2000; Ofer et al., 2017). Dies führt zu verschiedenen Szenarien über zeitliche Merkmale, die sich aus der Kollateralisierung ergeben.
Erstens kann es eine synchrone Aktivierung in den Tochterzweigen geben. Die auditorischen Hirnstammbahnen projizieren über ein einziges bifurkierendes Axon zu ipsi- und kontralateralen Zielen (kurze bzw. längere physische Bahnen). Die isochrone Übertragung wird durch unterschiedliche Myelinisierung und Axonkaliber der beiden Tochteräste erreicht (d. h. kleineres Kaliber und kürzere intermodale Längen ipsilateral; Seidl, 2014; Seidl und Rubel, 2016). Da die meisten verzweigten Axone im Vergleich zu den Hirnstamm-Hörbahnen ein größeres Gebiet abdecken und weniger gut definierte Funktionen erfüllen, sind die Daten für andere Systeme weitgehend unvollständig oder fehlen. Im Falle der Synchronizität könnte man jedoch vorhersagen, dass proximale Verzweigungen (d. h. die intrinsischen Kollateralen von Meynert- oder anderen Pyramidenzellen) anatomische Spezialisierungen aufweisen, die zu längeren Leitungszeiten führen, um die längeren Entfernungen der extrinsischen Kollateralen zu kompensieren und an diese anzupassen. Wie bereits erwähnt, scheint diese einfache Vorhersage nicht zuzutreffen. Weitere Untersuchungen erfordern die Entnahme von Stichproben aus identifizierten Axonen über große Entfernungen, was nicht einfach zu bewerkstelligen wäre.
Zweitens kann die astspezifische Aktivierung asynchron sein. Dies könnte auf einen selektiven Ausfall der Übertragung und/oder asynchrone Leitungszeiten über den Axonstrang zurückzuführen sein (Abbildung 2; Huguenard, 2000; Bucher, 2016). Modelle kortikaler Schaltkreise beschreiben ausgeprägte Routing-Zustände kurzzeitiger transienter Synchronie, die den Informationsfluss dynamisch gestalten könnten (Palmigiano et al., 2017). Für Langstreckenaxone fehlen vergleichbare experimentelle Daten weitgehend. Die kollaterale Topologie thalamokortikaler Projektionen ist jedoch ein Beispiel für ein kompliziertes, aktivitäts- und/oder zustandsabhängiges asynchrones Aktivierungsmuster.
Kortikale und thalamokortikale Aktivität ist in hohem Maße zustandsabhängig; und die Interaktion von präsynaptischen extrinsischen Eingängen (branchenspezifisch?) mit intrinsischen Membran- und synaptischen Eigenschaften postsynaptischer Neuronen wird als grundlegend für die Erzeugung rhythmischer Aktivität angesehen (mit „weitreichenden Effekten von der Verstärkung oder Blockierung sensorisch-motorischer Verarbeitung….“, McCormick et al, 2015).
Die Variabilität der kortikalen Antworten ist paradox, da diese auch als Substrate für stabile sensorische Erfahrungen dienen. Die neuronale Variabilität wurde mit dem Grad der heterogenen Synchronität über den extrinsischen Input in Verbindung gebracht; das heißt, dass sensorische Stimulationen oder Verhaltensbedingungen, die die Homogenität des Inputs in einem bestimmten Bereich erhöhen, auch die neuronale Variabilität verringern (Gomez-Laberge et al., 2016). Jüngste Diskussionen über die Reaktionen der Mikroschaltkreise haben über eine herausragende Rolle von leichten Abweichungen oder Unterschieden in der Information spekuliert: „Was aber, wenn die Unterschiede in der Konnektivität innerhalb von Kohorten von Zellen der gleichen Klasse für die Funktion des Schaltkreises wichtig sind?“ (Morgan und Lichtman, 2017).
Dynamische Axoneigenschaften
Änderungen in der Reaktionslatenz wurden im Zusammenhang mit verschiedenen Zuständen der Wachsamkeit berichtet. Im kortikothalamischen Signalweg führt eine erhöhte Wachsamkeit zu einer signifikant verkürzten Antwortlatenz. Dies und/oder Veränderungen in der Feuerungsfrequenz der ankommenden Impulse könnten für eine dramatisch erhöhte Antwortzuverlässigkeit der Teilpopulation (58 %) der visuell ansprechenden kortikogenikulären Neuronen verantwortlich sein (bei Kaninchen: Stoelzel et al., 2017). Diese Ergebnisse beziehen sich auf physiologisch identifizierte Einzelaxone; man kann jedoch über eine breitere Anwendbarkeit auf Verzweigungen kollateralisierter Axone spekulieren.
Fortlaufende Prozesse der Synaptogenese und des distalen Axonumsatzes wurden im erwachsenen Kortex nachgewiesen (NHP: Stettler et al., 2006). Auf kürzeren Zeitskalen zeigt die Superauflösungsmikroskopie von nicht-myelinisierten GFP-markierten CA3-Hippocampus in organotypischen Hirnschnitten, dass sich die Axone nach hochfrequenten Feuerstößen allmählich verbreitern, eine Beobachtung, die durch elektrophysiologische Aufzeichnungen bestätigt wird (Chereau et al., 2017). Andere, zweigspezifische Veränderungen werden wahrscheinlich noch entdeckt werden; zum Beispiel haben terminale Verzweigungen von individuell markierten Axonen aus der dorsalen Raphe einen zielspezifischen Prozentsatz von Boutonen, die das Protein VGLUT3 enthalten (größerer Prozentsatz für Zweige, die im Striatum enden als im motorischen Kortex). Dies deutet auf einen komplexen, uneinheitlichen Transportmechanismus über die Kollateralen hin (Gagnon und Parent, 2014).
Schlussfolgerung
In diesem Perspektivartikel habe ich die Axonverzweigung als relevant für Veränderungen des Gehirnzustands erörtert, wobei die Auswirkungen durch zweigspezifische Eigenschaften, unterschiedliche Rekrutierung postsynaptischer Ensembles und Synchronisationsmuster im gesamten Gehirn bewirkt werden. Dies knüpft an langjährige Diskussionen über axonale Verzweigungstopologien an und darüber, wie diese die Informationsverarbeitung durch Zeitverzögerungen bei der Impulsausbreitung, differentielle verzweigungsspezifische Filterung und aktivitätsabhängige Erregbarkeit modulieren könnten (z. B. Segev und Schneidman, 1999). Mit wenigen Ausnahmen, wie z. B. der auditorischen Hirnstammbahn, fehlen noch weitgehend harte Daten über synchrone und asynchrone Aktivierungen durch Tochterzweige und darüber, wie sich diese zeitlichen Beziehungen auf die postsynaptische neuronale Reaktionsfähigkeit auswirken könnten (siehe jedoch Gomez-Laberge et al., 2016; Stoelzel et al., 2017). Eine anhaltende Herausforderung besteht daher darin, verzweigungsspezifische Merkmale innerhalb einzelner Axone und die Auswirkungen auf postsynaptische Ensembles zu erforschen. Neuere Arbeiten werfen zusätzliche Fragen zur Netzwerkheterogenität auf, darunter auch, warum Neuronen aus einem einzigen Quellgebiet variabel zu einem oder mehreren Zielen projizieren, was immer wieder als „in allen Kombinationen“ beschrieben wird.
Autorenbeiträge
Der Autor bestätigt, dass er der einzige Autor dieser Arbeit ist und sie zur Veröffentlichung freigegeben hat.
Finanzierung
Wir danken den National Institutes of Health (NIH) für die finanzielle Unterstützung: MH107456.
Conflict of Interest Statement
Der Autor erklärt, dass die Forschung in Abwesenheit von kommerziellen oder finanziellen Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Barry, J. M. (2015). Axonale Aktivität in vivo: Technische Überlegungen und Implikationen für die Erforschung neuronaler Schaltkreise bei frei beweglichen Tieren. Front. Neurosci. 9:153 doi: 10.3389/fnins.2015.00153
PubMed Abstract | CrossRef Full Text | Google Scholar
Gagnon, D., and Parent, M. (2014). Distribution og VGLUT3 in highly collateralized axons from the rat dorsal raphe nucleus as revealed by single-neuron reconstructions. PLoS ONE 9:e87709. doi: 10.1371/journal.pone.0087709
CrossRef Full Text | Google Scholar
Kita, T., and Kita, H. (2012). Der Nucleus subthalamicus ist eine von mehreren Innervationsstellen für weitreichende kortikofugale Axone: eine Einzelaxon-Tracing-Studie bei der Ratte. J. Neurosci. 32, 5990-5999 doi: 10.1523/JNEUROSCI.5717-11
PubMed Abstract | CrossRef Full Text | Google Scholar