Mt Hood Community College Biology 102
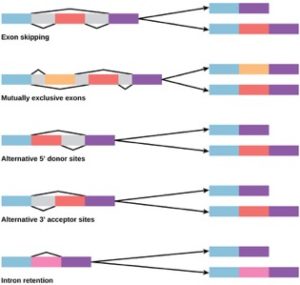
1970年代、alternative RNA splicingを示す遺伝子が初めて観察された。 代替RNAスプライシングは、イントロン(時にはエクソン)の異なる組み合わせが転写物から除去されたとき、1つの遺伝子から異なるタンパク質産物が生成されるようにするメカニズムである(図9.23)。 この代替スプライシングは無秩序に行われることもあるが、より多くの場合、制御され、遺伝子制御のメカニズムとして働く。異なる細胞や発生の異なる段階において異なるタンパク質産物の生産を制御する方法として、細胞によって異なるスプライシング代替物の頻度が制御されている。 ある推定によると、ヒトの遺伝子の 70% は、代替スプライシングによって複数のタンパク質として発現しています。
特に断りのない限り、このページの画像は OpenStax による CC-BY 4.0 の下でライセンス供与されています。 OpenStax CNX. 2016年5月18日http://cnx.org/contents/[email protected]
に掲載されました。