Amphicoelias
| Amphicoelias | |
|---|---|
| 240px | |
| Klasyfikacja naukowa | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Sauropsida |
| Klad: | Dinosauria |
| Nadrząd: | Sauropodomorpha |
| Rodzina: | †Diplodocidae |
| Genus: | †Amphicoelias Cope, 1878 |
| Gatunek typowy | |
| †Amphicoelias altus Cope, 1878a |
|
Amphicoelias (/ˌæmfᵻˈsiːliəs/, co oznacza „dwuwklęsły”, od greckiego αμφι, amphi: „po obu stronach”, i κοιλος, koilos: „pusty, wklęsły”) – rodzaj roślinożernych dinozaurów z rzędu zauropodów. Obejmuje to, co czasami było szacowane jako największy okaz dinozaura, jaki kiedykolwiek odkryto, pierwotnie nazwany „A. fragillimus”. Na podstawie zachowanych opisów pojedynczej kości kopalnej naukowcy przez lata szacowali, że A. fragillimus był najdłuższym znanym zwierzęciem o długości 58 metrów (190 stóp) i potencjalnie masie do 122,4 ton (134,9 krótkich ton). Jednak, ponieważ jedyne szczątki kopalne zostały utracone w pewnym momencie po ich zbadaniu i opisaniu w latach 70-tych XIX wieku, dowody przetrwały tylko w postaci rysunków i notatek terenowych. Nowsze analizy zachowanych dowodów oraz biologicznej wiarygodności tak dużego zwierzęcia lądowego sugerują, że ogromne rozmiary tego zwierzęcia były przeszacowane częściowo z powodu błędów typograficznych w oryginalnym opisie z 1878 roku. Według wielu naukowców dinozaur ten może nie istnieć.
Opis
Gatunek typu Amphicoelias, A. altus, został nazwany przez paleontologa Edwarda Drinkera Cope’a w grudniu 1877 r. (choć nie został opublikowany do 1878 r.) dla niekompletnego szkieletu składającego się z dwóch kręgów, kości łonowej (biodrowej) i kości udowej (górnej kości nogi). W tej samej pracy Cope nazwał także drugi gatunek, A. fragillimus. Jednak wszyscy późniejsi badacze uznali A. fragillimus za synonim A. altus. Już w 1881 roku uznano jednak, że A. altus nie da się odróżnić od innych rodzajów, gdyż cechy opisane przez Cope’a zostały źle zinterpretowane i są szeroko rozpowszechnione. W 1921 roku Osborn i Mook przypisali dodatkowe kości do A. altus – łopatkę (shoulder blade), kość ramienną (coracoid), kość łokciową (lower arm bone) i ząb. Henry Fairfield Osborn i Charles Craig Mook zauważyli ogólne podobieństwo między Amphicoelias i Diplodocus, jak również kilka kluczowych różnic, takich jak proporcjonalnie dłuższe kończyny przednie u Amphicoelias niż u Diplodocus. Uzębienie Amphicoelias jest homodontyczne. Jego zęby mają kształt długich, smukłych, cylindrycznych prętów, są rozstawione i wystają do przodu, w kierunku przedniej części pyska. Kość udowa Amphicoelias jest niezwykle długa, smukła i okrągła w przekroju; choć kiedyś uważano, że ta okrągłość jest kolejną cechą wyróżniającą Amphicoelias, od tego czasu znaleziono ją także u niektórych okazów Diplodocus. A. altus był również podobnej wielkości co Diplodocus, szacowany na około 25 metrów (82 stopy) długości. Podczas gdy większość naukowców użyła tych szczegółów, by odróżnić Amphicoelias i Diplodocus jako oddzielne rodzaje, przynajmniej jeden zasugerował, że Amphicoelias jest prawdopodobnie starszym synonimem Diplodocus.
Historia
Amphicoelias fragillimus został zebrany przez Oramela Lucasa, kolekcjonera skamieniałości zatrudnionego przez E. D. Cope’a, wkrótce po tym, jak został zatrudniony przez Cope’a w 1877 roku. Lucas odkrył częściowy kręg (łuk neuralny i kręgosłup) nowego gatunku zauropoda w Garden Park, na północ od Cañon City w Kolorado, w pobliżu kamieniołomu, z którego pochodzi Camarasaurus. Kręg był w złym stanie, ale zadziwiająco duży, mierzył od 1,5 metra (4,9 stopy) do 2,7 metra (8,9 stopy) wysokości. Lucas wysłał ten okaz do Cope’a wiosną lub wczesnym latem 1878 roku, a Cope opublikował go jako holotyp (numer katalogowy AMNH 5777) nowego gatunku, A. fragillimus, w sierpniu tego samego roku. Nazwa pochodzi od łacińskiego fragillimus („bardzo kruchy”), odnoszącego się do delikatności kości wytwarzanej przez bardzo cienkie blaszki (ściany kręgów). Jak wynika z notatników Cope’a, które zapisał na podstawie raportu Lucasa o lokalizacji wykopalisk w 1879 roku, okaz pochodził ze wzgórza na południe od kamieniołomu Camarasaurus, znanego obecnie jako „Cope’s Nipple”. Podczas gdy Cope pierwotnie napisał, że miejsce to należało do formacji Dakota (środkowa kreda w wieku), obecność dinozaurów takich jak Camarasaurus w tych samych skałach wskazuje, że prawdopodobnie należą one do formacji Morrison, co umieszcza wiek miejsca na 150 milionów lat temu w późnej Jurze, a konkretnie w wieku Tithonian.
Gigantyczne kości przypisywane A. fragillimus były często ignorowane w podsumowaniach największych dinozaurów częściowo dlatego, że według różnych późniejszych doniesień miejsce pobytu zarówno kręgów, jak i kości udowej jest nieznane, a wszystkie próby ich zlokalizowania zakończyły się niepowodzeniem. Carpenter w 2006 roku przedstawił możliwy scenariusz zaginięcia okazów A. fragillimus. Jak zauważył Cope w swoim opisie, materiał kostny z łuku neuralnego był bardzo kruchy, a techniki utwardzania i konserwacji kości kopalnych nie zostały jeszcze wynalezione (rywal Cope’a, paleontolog O.C. Marsh, jako pierwszy użył takich chemikaliów na początku lat osiemdziesiątych XIX wieku). Carpenter zauważył, że skamieniałe kości znane z kamieniołomu A. fragillimus byłyby zachowane w głęboko zwietrzałym mułowcu, który ma tendencję do łatwego kruszenia się i fragmentacji na małe, nieregularne kostki. Dlatego kość mogła się źle pokruszyć i zostać wyrzucona przez kogoś w Amerykańskim Muzeum Historii Naturalnej (być może nawet przez samego Cope’a) wkrótce po tym, jak zilustrował ją w widoku od tyłu do swojej pracy. Carpenter zasugerował, że może to wyjaśniać, dlaczego Cope narysował ten kręg tylko w jednym widoku, a nie pod wieloma kątami, jak to zrobił w przypadku innych swoich odkryć.
W 1994 roku podjęto próbę przeniesienia oryginalnego kamieniołomu, w którym znaleziono A. fragillimus i inne gatunki, używając radaru penetrującego grunt w próbie zobrazowania kości wciąż zakopanych w ziemi. Próba ta nie powiodła się, ponieważ skamieniałe kości z mułowca miały taką samą gęstość jak otaczająca je skała, co uniemożliwiło ich rozróżnienie. Badanie lokalnej topografii wykazało również, że warstwy skamieniałych skał były mocno zerodowane i prawdopodobnie było tak w czasie, gdy Lucas odkrył A. fragillimus, wskazując, że większość szkieletu zniknęła do czasu, gdy odzyskano kręgi i kości udowe.
W 1884 r. opis Cope’a dotyczący A. fragillimus spotkał się ze sceptycyzmem, a niektórzy badacze zauważyli błędy typograficzne w jego pomiarach. Na przykład, jednostki pomiarowe podane są w (oczywiście niepoprawnych) centymetrach, a nie milimetrach. Carpenter argumentował, że są wszelkie powody, by wierzyć Cope’owi na słowo, zauważając, że stawką była reputacja paleontologa. Odkrycie miało miejsce w czasie Wojen o Kości, a rywal Cope’a, Marsh, który był „zawsze gotów upokorzyć” Cope’a, nigdy nie zakwestionował jego twierdzeń. Marsh był znany z tego, że zatrudniał szpiegów do monitorowania odkryć Cope’a, a nawet mógł mieć potwierdzenie ogromnych rozmiarów kości Amphicoelias fragillimus. Paleontolodzy Henry Fairfield Osborn i C.C. Mook w 1921 roku, a także John S. McIntosh w 1998 roku, również zaakceptowali dane Cope’a bez wątpliwości w opublikowanych recenzjach. W 2015 roku Woodruff i Foster opublikowali analizę dowodów i okoliczności towarzyszących publikacji i interpretacji A. fragillimus.
Rozmiar
Oszacowanie kompletnego rozmiaru A. fragillimus wymaga przeskalowania kości lepiej znanych gatunków diplodocidów (rodzina niezwykle długich i smukłych zauropodów) przy założeniu, że ich względne proporcje były podobne. W swojej oryginalnej pracy Cope dokonał tego, spekulując na temat wielkości hipotetycznej kości udowej (górnej części nogi) A. fragillimus. Cope zauważył, że u innych zauropodów, zwłaszcza A. altus i Camarasaurus supremus, kości udowe były zawsze dwa razy wyższe niż najwyższy kręg grzbietowy, i oszacował rozmiar kości udowej A. fragillimus na 12 stóp (3,6 m) wysokości.
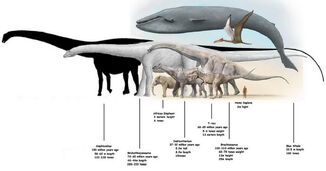
W 1994 r., używając pokrewnego Diplodocus jako odniesienia, Gregory S. Paul oszacował długość kości udowej A. fragillimus na 3,1 do 4 metrów (10 do 13 stóp). Ken Carpenter, który w 2006 roku dokonał ponownej oceny A. fragillimus, również użył Diplodocus jako wzorca, ustalając wysokość kości udowej na 4,3 do 4,6 m (14 do 15 stóp). Carpenter oszacował kompletny rozmiar A. fragillimus, choć zastrzegł, że względne proporcje u diplodoków mogą się różnić w zależności od gatunku. Przyjmując te same proporcje co u dobrze znanego Diplodocus, Carpenter przedstawił szacunkową długość całkowitą 58 m (190 stóp), która, jak zauważył, mieściła się w zakresie przedstawionym przez Paula w 1994 r. (40-60 metrów (130-200 stóp)). Carpenter zaznaczył, że nawet najniższe szacunki długości A. fragillimus były wyższe niż te dla innych olbrzymich zauropodów, takich jak diplodok Supersaurus (32,5 m), brachiozauryd Sauroposeidon (34 m) i tytanozaur Argentinosaurus (30 m). Carpenter przedstawił bardziej spekulatywne, konkretne proporcje dla A. fragillimus (znów na podstawie przeskalowanego Diplodocus), w tym długość szyi 16,75 m (55 stóp), długość ciała 9,25 m (30,3 stóp) i długość ogona 32 m (105 stóp). Całkowitą wysokość kończyn przednich oszacował na 5,75 metra (18,9 ft), wysokość kończyn tylnych na 7,5 metra (25 ft), a wysokość całkowitą (w najwyższym punkcie grzbietu) na 9,25 metra (30,3 ft). Dla porównania wieloryb błękitny, który jest średnio najdłuższym żyjącym stworzeniem, osiąga 29,9 metra (98 stóp) długości.
While A. fragillimus był stosunkowo cienki, jego ogromny rozmiar wciąż czynił go bardzo masywnym. Waga jest znacznie trudniejsza do określenia niż długość u zauropodów, gdyż bardziej złożone równania są podatne na większe marginesy błędu oparte na mniejszych różnicach w ogólnych proporcjach zwierzęcia. Carpenter wykorzystał oszacowaną przez Paula w 1994 r. masę Diplodocus carnegii (11,5 tony metrycznej (11,3 tony długiej; 12,7 tony krótkiej)), by spekulować, że A. fragillimus mógł ważyć nawet 122,4 tony metrycznej (120,5 tony długiej; 134,9 tony krótkiej). Najcięższy zarejestrowany wieloryb błękitny ważył 173 tony metryczne (170 ton długich; 191 ton krótkich), a najcięższy dinozaur znany z dość dobrych szczątków, Argentinosaurus, ważył od 80 do 100 ton metrycznych (79 do 98 ton długich; 88 do 110 ton krótkich), choć jeśli szacunki rozmiarów uda się potwierdzić, mógł być jeszcze lżejszy od Bruhathkayosaurus, którego wagę oszacowano na 126 ton metrycznych (124 tony długie; 139 ton krótkich), ale znany jest też z bardzo fragmentarycznych szczątków.
Klasyfikacja i gatunki
Edward Drinker Cope opisał swoje znaleziska w dwóch wydaniach „American Naturalist” z 1878 roku i przypisał je do nowego rodzaju Amphicoelias. Umieścił go w unikalnej rodzinie Amphicoeliidae, choć jest to obecnie uważane za nomen oblitum (zapomnianą nazwę). Rodzaj jest zwykle przypisywany do rodziny Diplodocidae, choć niektóre współczesne analizy wykazały, że znajduje się on u podstawy większej grupy Diplodocoidea lub jako diplodokoid incertae sedis (niepewne umiejscowienie). Pierwszy nazwany gatunek w rodzaju, Amphicoelias altus (okaz holotypowy AMHD 5764), został odkryty przez Cope’a w 1877 roku. Chociaż jest on reprezentowany tylko przez częściowy szkielet, to jednak istnieje wystarczająco dużo cech diagnostycznych, by wstępnie zdefiniować rodzaj. A. altus znany jest z lepszych szczątków, ale jest mniejszy niż A. fragillimus. Cope w 1878 roku nazwał także drugi gatunek: Amphicoelias latus.
Trzeci nazwany gatunek Amphicoelias, A. fragillimus, znany był tylko z pojedynczego, niekompletnego 1,5-metrowego łuku neuralnego (część kręgu z kolcami i wyrostkami), ostatniego lub przedostatniego w serii kręgów grzbietowych, D (dorsal) 10 lub D9. Opierając się jedynie na ilustracji opublikowanej w 1878 roku, kręg ten mierzyłby w życiu 2,7 metra (8,9 stopy) wysokości. Twierdzi się jednak, że pasek skali w opublikowanym opisie zawierał błąd typograficzny, a kopalny kręg miał w rzeczywistości tylko 1,38 metra (4,5 stopy) wysokości. Oprócz tego kręgu, notatki terenowe Cope’a zawierają wpis o „mmense distal end of femur”, znajdującym się zaledwie kilkadziesiąt metrów od olbrzymiego kręgu. Jest prawdopodobne, że ta nieopisana kość nogi należała do tego samego osobnika, co kręgosłup neuralny.
W 2010 roku udostępniono monografię, ale nie została ona formalnie opublikowana, autorstwa Henry’ego Galiano i Raimunda Albersdorfera, w której odnieśli oni czwarty gatunek do Amphicoelias, jako „A. brontodiplodocus” na podstawie kilku kompletnych okazów znalezionych w kamieniołomie Dana w Big Horn Basin w stanie Wyoming i przechowywanych w prywatnej kolekcji. Ta specyficzna nazwa odnosiła się do ich hipotezy opartej na tych okazach, że prawie wszystkie gatunki diplodoków z Morrison są albo stadiami rozwojowymi, albo reprezentują dymorfizm płciowy wśród członków rodzaju Amphicoelias, ale ta analiza spotkała się ze sceptycyzmem, a sama publikacja została zdyskwalifikowana przez jej głównego autora, wyjaśniającego, że jest to „oczywiście zredagowany manuskrypt z literówkami, etc., a nie ostateczna praca”. In fact, no printing or distribution has been attempted”.
Osborn i Mook w 1921 r. dokonali tymczasowej synonimizacji trzech gatunków, zatapiając A. latus w Camarasaurus supremus, sugerując jednocześnie, że A. fragillimus to po prostu bardzo duży osobnik A. altus, z którym to stanowiskiem zgodziła się większość późniejszych badań, w tym McIntosh 1998, Foster (2007) oraz Woodruff i Foster (2015). Carpenter (2006) nie zgodził się jednak co do synonimiczności A. altus i A. fragillimus, powołując się na liczne różnice w budowie kręgów zauważone także przez Cope’a, i zasugerował, że różnice te są wystarczające, by uzasadnić osobny gatunek lub nawet osobny rodzaj dla A. fragillimus. Ostrzegł jednak, że uznanie A. fragillimus za odrębny gatunek jest prawie niemożliwe do ustalenia bez oryginalnego okazu do badań. Choć Amphicoelias latus wyraźnie nie jest Amphicoelias, to prawdopodobnie jest synonimem Camarasaurus grandis, a nie C. supremus, ponieważ został znaleziony niżej w formacji Morrison, a głęboko wklęsłe powierzchnie stawowe na kręgach ogonowych są bardziej spójne z C. grandis.
W 2007 roku John Foster zasugerował, że różnice zwykle przytaczane w celu odróżnienia Amphicoelias altus od bardziej znanego Diplodocus nie są znaczące i mogą wynikać z indywidualnej zmienności. Foster argumentował, że Amphicoelias jest prawdopodobnie starszym synonimem Diplodocus i że jeśli dalsze badania to potwierdzą, trzeba będzie porzucić znaną nazwę Diplodocus na rzecz Amphicoelias, tak jak to było w przypadku Brontosaurus i jego starszego synonimu Apatosaurus. W 2015 roku Woodruff i Foster powtórzyli ten wniosek, stwierdzając, że istnieje tylko jeden gatunek Amphicoelias i że można go odnieść do Diplodocus jako Diplodocus altus. Uznali oni nazwę Amphicoelias za nomen oblitum. Wysunięto też hipotezę, że Amphicoelias powinien być uznany za apatozaura, a zatem powinien być umieszczony w podrodzinie Apatosaurinae.
Następujący kladogram Diplodocidae za Tschoppem, Mateusem i Bensonem (2015) zamiast tego pokazuje A. altus poza Diplodocinae.
Diplodocidae
Amphicoelias altus
Apatosaurinae
Gatunek nienazwany
Apatosaurus ajax
Apatosaurus louisae
Brontosaurus excelsus
Brontosaurus yahnahpin
Brontosaurus parvus
Diplodocinae
Gatunek nienazwany
Tornieria africana
.
Supersaurus lourinhanensis
Supersaurus vivianae
Leinkupal laticauda
Galeamopus hayi
.
Diplodocus carnegii
Diplodocus hallorum
Kaatedocus siberi
Barosaurus lentus
Paleobiologia
W swojej ponownej w 2006 r.ocenie, Carpenter zbadał paleobiologię olbrzymich zauropodów, w tym Amphicoelias, i odniósł się do pytania, dlaczego ta grupa osiągnęła tak ogromne rozmiary. Wskazał, że gigantyczne rozmiary zostały osiągnięte wcześnie w ewolucji zauropodów, z bardzo dużymi gatunkami obecnymi już w późnym triasie, i stwierdził, że jakakolwiek presja ewolucyjna powodująca duże rozmiary była obecna od wczesnych początków grupy. Carpenter przytoczył kilka badań nad olbrzymimi ssakami roślinożernymi, takimi jak słonie i nosorożce, które wykazały, że większy rozmiar u zwierząt roślinożernych prowadzi do większej wydajności w trawieniu pokarmu. Ponieważ większe zwierzęta mają dłuższe układy pokarmowe, pokarm jest trawiony przez znacznie dłuższy czas, co pozwala dużym zwierzętom przetrwać na gorszej jakości źródłach pokarmu. Jest to szczególnie prawdziwe w przypadku zwierząt z dużą liczbą „komór fermentacyjnych” wzdłuż jelita, które pozwalają mikrobom gromadzić się i fermentować materiał roślinny, wspomagając trawienie. W całej swojej historii ewolucyjnej zauropody występowały głównie w półpustynnych, sezonowo suchych środowiskach, z odpowiednim sezonowym spadkiem jakości pożywienia w porze suchej. Środowisko Amphicoelias było zasadniczo sawanną, podobną do jałowych środowisk, w których żyją współczesne olbrzymie zwierzęta roślinożerne, co potwierdza tezę, że słabej jakości pożywienie w jałowym środowisku sprzyja ewolucji olbrzymich roślinożerców. Carpenter argumentował, że inne korzyści z dużych rozmiarów, takie jak względna odporność na drapieżniki, niższe wydatki energetyczne i dłuższe życie, są prawdopodobnie zaletami drugorzędnymi.
Środowisko formacji Morrison, w którym żył Amphicoelias, przypominałoby współczesną sawannę, choć ponieważ trawa nie pojawiła się aż do późnej kredy, paprocie były prawdopodobnie dominującą rośliną i głównym źródłem pożywienia dla Amphicoelias. Choć Engelmann i in. (2004) odrzucili paprocie jako źródło pożywienia zauropodów ze względu na ich stosunkowo niską kaloryczność, Carpenter argumentował, że układ pokarmowy zauropodów, dobrze przystosowany do radzenia sobie z pokarmem niskiej jakości, pozwala na spożywanie paproci jako dużej części diety zauropodów. Carpenter zauważył również, że sporadyczna obecność dużych skamieniałych kłód wskazuje na obecność wysokich na 20-30 m (66-98 stóp) drzew, co wydawałoby się sprzeczne z porównaniem z sawanną. Drzewa te są jednak rzadkie, a ponieważ wysokie drzewa wymagają więcej wody niż środowisko sawanny mogło na ogół zapewnić, prawdopodobnie istniały w wąskich traktach lub „lasach galeriowych” wzdłuż rzek i wąwozów, gdzie woda mogła się gromadzić. Carpenter spekulował, że gigantyczne zwierzęta roślinożerne, takie jak Amphicoelias, mogły wykorzystywać cień lasów galeriowych, by zachować chłód w ciągu dnia, a w nocy robić większość pożywienia na otwartej sawannie.
Media
Zauropod, który może być potencjalnie zidentyfikowany jako Amphicoelias fragillimus, pojawia się w odcinku serialu telewizyjnego The Land Before Time, „The Hidden Canyon”. Identyfikacja wynika z faktu, że podczas gdy rozmiar normalnego dorosłego Land Before Time „Longneck”, używa modelu zwykle używanego tylko dla młodych postaci, takich jak Littlefoot i Ali. Może to potencjalnie oznaczać, że osobnik ten reprezentuje młodego osobnika jeszcze większego dinozaura; prawdopodobnie behemota A. fragillimus opisanego przez Cope’a w 1870 roku.
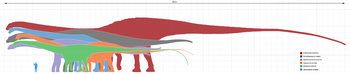
Amphicoelias był największym i najdłuższym dinozaurem
Galeria
Amphicoelias/Galeria
.