Frontiers in Systems Neuroscience
Changes in brain state are associated with microcircuitry changes in neuronal firing properties and with macro-level changes in synchronous or asynchronous patterns of brain activation. Neste contexto, os axônios foram relativamente menos investigados (Barry, 2015), e então principalmente no contexto da velocidade de condução dos potenciais de ação e do aumento da atenção que pode resultar de mudanças na velocidade de condução (por exemplo, Stoelzel et al., 2017). Além disso, entretanto, a topologia altamente ramificada de muitos axônios parece bem desenhada para recrutar grupos neuronais pós-sinápticos distribuídos, possivelmente de forma dependente do estado. O processamento de sinais pode ser efetuado em pelo menos três domínios axonais, a saber, mapeamento, amplificação e tempo (Innocenti et al., 2016; Innocenti, 2017). Nesta perspectiva, discuto brevemente, primeiro, exemplos de rede de colateralização axonal, e depois algumas das características axon-intrínsecas que podem estar subjacentes ao recrutamento pós-sináptico diferencial. Na ausência de parâmetros ou mecanismos detalhados, o objetivo é principalmente destacar características gerais que possam figurar no controle e transições dos estados cerebrais.
Axon Collaterals
Todos os axônios possuem uma elaborada arborização distal na estrutura-alvo. Muitos axônios, além disso, possuem múltiplos ramos (aka, colaterais) que têm como alvo estruturas distintas (revisado em Rockland, 2013, 2018). Um exemplo principal é a ramificação de axônios da camada 5 de neurônios corticotalâmicos. Isto tem sido documentado repetidamente e é frequentemente discutido como um mecanismo chave pelo qual um organismo pode distinguir se as mudanças no input sensorial são produzidas por mudanças no ambiente ou por movimentos auto-iniciados (“efference copy” ou “descarga colateral”, Guillery e Sherman, 2011).
Em roedores, onde há um grande número de técnicas aplicáveis para investigar esta questão, a colateralização é conhecida por ser relativamente comum. Para dar vários exemplos: (1) neurônios no projeto do subiculum hipocampal para os corpos mamilares, o córtex retroesplenial, ou por colaterais para ambos (rato: Kinnavane et al., 2018); (2) neurônios serotonérgicos no contato do núcleo do ráquis dorsal (DR) em várias combinações de estriato, córtex pré-frontal e amígdala (rato: Gagnon e Parent, 2014), bem como combinações de núcleos autonômicos (Waselus et al., 2011). Neste caso, a ativação da resposta ao estresse por colaterais DR pode alcançar a ativação sincronizada dos núcleos associados à liberação de neurohormona ou respostas pressóricas. As ativações diferenciais e coordenadas das estruturas do cérebro podem contribuir para as funções multifacetadas mas relacionadas dos DR, tais como regulação do ciclo sono-vigília, modulação dos sinais de dor, ou expressão do humor (Gagnon e Parent, 2014).
Terceiro estudo anatômico de projeções talamocorticais do rato identifica “eixos multiespecíficos” que se ramificam amplamente para domínios restritos em áreas corticais (e subcorticais) separadas. Estes têm sido hipotéticos para orquestrar a rápida emergência e reconfiguração de conjuntos neurais sincronizáveis e espacialmente distribuídos (Clasca et al., 2016).
Quarto, um estudo recente de conexões cortical-corticais usando o traçado axonal de todo o cérebro no córtex visual do mouse descobriu que 23 de 30 neurônios contataram de duas a sete outras áreas corticais. Em confirmação deste resultado, o seqüenciamento de DNA de alta produção de neurônios com código de barras genético encontrou 44% dos 533 neurônios a serem multiplicados projetando (“neurônios broadcast”, Han et al., 2018). Han et al. distinguiram provisoriamente dois tipos amplos de neurônios projetando, uma subpopulação menor “dedicada” (unidirecional), coexistindo com uma prevalência de células “broadcast” (projetando múltiplas). Poderia esta arquitetura subservir modulações em estado cognitivo e processamento sensorial?
Colateralização não estereotipada
Como observado acima, neurônios que colateralizam o fazem em um padrão não estereotipado. Dentro de uma projeção designada (definida pela origem), os neurônios enviam ramos para um subconjunto diversificado de áreas-alvo (“em todas as combinações”). Esta observação é tão consistente que pode ser considerada uma regra, embora os parâmetros detalhados não tenham sido tabulados. O significado funcional também não é claro, mas os resultados emergentes sugerem que os inputs heterogêneos e variáveis para um neurônio cortical (e, por extensão, podemos inferir para as assemblages neurais) são importantes na condução da variabilidade e mudanças de picos de trem através de experimentos experimentais (Gomez-Laberge et al, 2016 e veja abaixo).
Intrinsic Collaterals
Em todas as espécies, os neurônios de projeção cortical de longa distância, além de alvos extrínsecos simples ou múltiplos, normalmente têm uma arborização intrínseca elaborada. Alguns neurônios corticais têm apenas uma colateralização intrínseca (local), e alguns apenas uma extrínseca (rato: Kita e Kita, 2012). Embora existam poucos dados para as proporções reais de arborizações intrínsecas e extrínsecas (Parent et al., 2000; Rockland, 2018), as evidências sugerem que isso será altamente variável. Mesmo dentro do sistema de conexões intrínsecas, não somente existem diferenças no número de colaterais e número de boutons sinápticos, mas um único neurônio pode ter uma mistura de colaterais mielinizados e não mielinizados (cat visual cortex: Martin et al., 2014; Koestinger et al., 2017). A mielinização específica do ramo poderia resultar em aumento da velocidade de condução específica do ramo, embora Koestinger et al. sugiram que ela pode ter mais a ver com fatores como aumento da segurança de transmissão, presumivelmente, novamente, específicos do ramo.
Uma observação curiosa relacionada à mielinização diz respeito à estria de Gennari, a faixa mielinizada de axônios na camada 4B da área de primatas V1. Como esta consiste em colaterais intrínsecos, a explicação comum, de que a mielinização é um meio de aumentar a velocidade de condução em longas distâncias, não é imediatamente aplicável. Os colaterais locais não necessitam de (?) aumento em relação a alvos distantes e, de facto, para a sincronicidade que se poderia ter previsto, o aumento da condução local (via mielinização?) parece paradoxal. Poderia ser que a mielinização esteja relacionada a outros fatores, tais como mudanças relacionadas à plasticidade (ou ao estado) no diâmetro do axônio?
Um exemplo interessante: Células Meynert na Área V1 de Primatas Não Humanos
Estes grandes neurônios, na borda das camadas 5 e 6, projetam-se variavelmente para extrastriar a área MT, e/ou outras áreas de associação visual, e/ou para o núcleo pulvinar, e/ou para o colículo superior e o pretectum (Weisenhorn et al., 1995; Rockland e Knutson, 2001). A colateralização intrínseca é excepcionalmente extensa, medida a 8,0 mm do soma, com base nas reconstruções subtotais, e com pelo menos 800-1.370 boutons (Figura 1). Através da arborização axonal, parece haver uma variabilidade distinta no diâmetro dos ramos; isto é, os ramos extrínsecos dirigidos à área MT são grandes (diâmetro ~3,0 μm), mas os ramos intrínsecos e os que se projetam para o colículo pulvinar e superior parecem menores, a julgar pela microscopia da luz (cf. Figura 1B (intrínseca) e Figura 1D (extrínseca à área MT). Diâmetros variáveis presumivelmente indicam diferenças no grau de mielinização e, por inferência, na velocidade de condução.
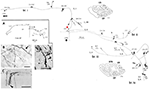
Figura 1. Uma típica arborização axonal proximal, espacialmente estendida, de uma célula de Meynert (asterisco vermelho) no córtex visual primário de um macaco macaco. Existem três principais colaterais intrínsecos (rotulados como br. i, ii, iv), estendendo 3,0 mm dorsal na camada 1, 3,0 mm ventral na camada 6, 4,0 mm ventral na camada 6, e 0,5 mm na camada 4B, como indicado pelas pontas de seta nos dois contornos das seções coronais (seções 222 e 274, onde dorsal é à esquerda). Todos juntos, os colaterais intrínsecos se estendem por 5,9 mm anterior-posterior (117 seções × 50 μm). Um outro colateral extrínseco (br. iii) ocorre nas camadas 4 e 6 da área V2. Porções dos colaterais individuais e do axônio principal (seta grossa) não puderam ser seguidas, como é indicado por linhas tracejadas. O inset de baixa ampliação (A, à esquerda) fornece uma visão esquemática da configuração geral. Os números denotam seções individuais, onde 20 números = 1,0 mm. Todos os ramos têm numerosos pequenos clusters sinápticos, um dos quais é ilustrado em (B). Observe o diâmetro diminuído entre o axônio principal (seta) e a arborização terminal. Os eixos extrínsecos (C) são de diâmetros variáveis (um eixo de grande diâmetro na ponta da flecha). (D) Terminações extrínsecas na área MT incluem alguns axônios de grande diâmetro. Barra de escala = 25 μm em (B), 100 μm em (C), 20μm em (D). CF, fissura calcarina; LS, sulco lunar; STS, sulco temporal superior; L, camada. Modificado das Figuras 1, 9 em Rockland e Knutson (2001) e Figuras 6f, 7b de Rockland (1995) com permissão.
Importância funcional da ramificação Axon
Um componente importante da colateralização é que os ramos filhas muitas vezes não são uniformes, mas especialmente em pontos de ramos, variam em diâmetro (Figura 2). A variabilidade no diâmetro juntamente com outros parâmetros terá impacto na excitabilidade, velocidade de condução e outros aspectos da propagação do sinal. Outros parâmetros de impacto incluem largura da mielina e comprimento intermodal, e densidade e distribuição dos canais iônicos (revisado em Debanne et al., 2011; Seidl, 2014; Bucher, 2016; Seidl e Rubel, 2016; Rama et al., 2018). Estes teriam efeitos nas propriedades de resposta neural ao nível do microcircuito. A nível mais global, a atividade somada de múltiplos eixos de projeção, com velocidades de condução variáveis através de uma rede interligada, poderia resultar em um espectro de ativações síncronas e/ou assíncronas (Mitra et al., 2015; Zeki, 2016). O recrutamento diferencial de populações pós-sinápticas ou recombinações da rede poderiam ser fatores em transições de estado ou modulação.
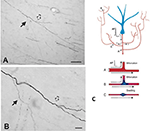
Figura 2. Um segmento tipicamente ramificado de axônio na matéria branca (macaco macaco). O segmento é originário de um neurônio no córtex parietal e é visto aqui na vizinhança do córtex temporal ventral. O painel (A) tem uma ampliação inferior a (B). Note bifurcações duplas, onde o primeiro ramo filho (seta sólida) é visivelmente mais fino (e não mielinizado?) do que o axônio principal. Na segunda, ligeiramente mais distal bifurcação (seta oca), os ramos filhas aparecem de diâmetro aproximadamente igual, mas ambos são mais finos do que o axônio principal. Barras de escala = 100 μm (A) e 10 μm (B). Reproduzido de Zhong e Rockland (2003) com permissão. (C) Esquema de um neurônio (azul) e sua topologia de ramificação estendida (pre-fechada para uma formatação conveniente). Um potencial de ação (AP) pode seguir caminhos sinuosos (A-C) para múltiplos alvos. A confiabilidade de propagação depende tanto das propriedades elétricas ativas do axônio quanto de sua geometria, incluindo as não homogeneidades da membrana, tais como inchaços e diâmetros incompatíveis de ramificações. Abaixo: esquema para ilustrar a propagação confiável (A, com ótima correspondência de impedância entre os ramos mãe e filha), e propagação lenta ou fracassada (B, onde o ramo filha tem um diâmetro aumentado; C, onde há um inchaço da membrana interposta). Reproduzido de Huguenard (2000) com permissão.
A geometria do axônio, as propriedades elétricas ativas e a não homogeneidade da membrana em pontos de ramos são bem conhecidas como fatores de propagação confiáveis (ex., Manor et al., 1991; Innocenti et al., 1994; Tettoni et al., 1998; Huguenard, 2000; Ofer et al., 2017). Isto leva a vários cenários diferentes sobre características temporais conseqüentes à colateralização.
Primeiro, pode haver ativação síncrona em todos os ramos filhas. As vias do tronco cerebral auditivo projetam-se através de um único eixo bifurcante para alvos ipsi e contralaterais (respectivamente, vias físicas curtas e longas). A transmissão isocrônica é alcançada por mielinização diferencial e calibre axonal das duas ramificações filhas (ou seja, calibre menor e comprimentos intermodais mais curtos ipsilateralmente; Seidl, 2014; Seidl e Rubel, 2016). Como a maioria dos axônios ramificados, em comparação com as vias auditivas do tronco cerebral, cobre um território maior e sub-reserva funções menos bem definidas, os dados são em grande parte incompletos ou inexistentes para outros sistemas. Pode-se prever, entretanto, no caso de sincronicidade, que os ramos proximais (ou seja, os colaterais intrínsecos de Meynert ou outras células piramidais) teriam especializações anatômicas resultando em tempos de condução mais longos, para compensar e combinar com as distâncias mais longas dos colaterais extrínsecos. Como já foi referido, esta simples previsão não parece ser válida. Uma investigação mais profunda implicaria na amostragem de eixos identificados em longas distâncias, e não seria fácil de se obter.
Segundo, a ativação específica do ramo pode ser assíncrona. Isto poderia ser devido a falha seletiva de transmissão e/ou tempos de condução assíncronos através do eixo axonal (Figura 2; Huguenard, 2000; Bucher, 2016). Modelos de circuitos corticais descrevem estados distintos de roteamento de sincronia transitória de curta duração que poderiam moldar dinamicamente o fluxo de informação (Palmigiano et al., 2017). Faltam dados experimentais comparáveis para os axônios de longa distância. A topologia colateral das projeções talamocóricas, entretanto, fornece um exemplo evocativo de um padrão de ativação assíncrona complicado, dependente de atividade e/ou estado.
Atividade cortical e talamocórtica é altamente dependente de estado; e a interação de entradas extrínsecas pré-sinápticas (específicas do ramo?) com propriedades intrínsecas de membrana e sinápticas dos neurônios pós-sinápticos é considerada fundamental para a geração de atividade rítmica (com “efeitos que vão desde o aumento ou bloqueio do processamento sensório-motor….”, McCormick et al, 2015).
Variabilidade nas respostas corticais é paradoxal uma vez que estas também servem como substratos de experiência sensorial estável. A variabilidade neuronal tem sido associada com o grau de sincronia heterogênea através do input extrínseco; ou seja, a estimulação sensorial ou condições comportamentais que aumentam a homogeneidade de input para uma determinada área é prevista para também reduzir a variabilidade neuronal (Gomez-Laberge et al., 2016). Discussões recentes sobre respostas de microcircuitos têm especulado sobre um papel proeminente de pequenas variações ou diferenças na informação: “Mas e se as diferenças entre a conectividade dentro de coortes de células da mesma classe forem importantes para o funcionamento do circuito?” (Morgan e Lichtman, 2017).
Propriedades Dinâmicas do Axon
Alterações na latência de resposta foram relatadas em relação a diferentes estados de alerta. Na via corticotalâmica, o aumento da atenção resulta em uma latência de resposta significativamente reduzida. Isto e/ou mudanças na freqüência de disparo dos impulsos de chegada podem ser responsáveis por um aumento dramático da confiabilidade de resposta para a subpopulação (58%) de neurônios corticogeniculados visualmente sensíveis (em coelhos: Stoelzel et al., 2017). Estes resultados dizem respeito aos axônios únicos fisiologicamente identificados; mas pode-se especular sobre uma aplicabilidade mais ampla a ramos de axônios colateralizados.
Processos contínuos de sinaptogênese e rotação distal de axônios têm sido demonstrados no córtex adulto (NHP: Stettler et al., 2006). Em escalas de tempo mais curtas, a microscopia de super-resolução do hipocampo CA3 não mielinizado em cortes organotípicos do cérebro mostra que os axônios se alargam gradativamente após episódios de queima de alta freqüência, uma observação confirmada pelo registro eletrofisiológico (Chereau et al., 2017). Outras mudanças, específicas dos ramos são prováveis de serem descobertas; por exemplo, arborizações terminais de axônios rotulados individualmente a partir do ráquis dorsal têm uma porcentagem específica de alvos de boutons que contêm a proteína VGLUT3 (maior porcentagem para ramos que terminam no estriato do que no córtex motor). Isto implica um mecanismo complexo e não uniforme de tráfico de colaterais (Gagnon e Parent, 2014).
Conclusão
Neste artigo Perspectiva, discuti a ramificação dos axônios como sendo relevante para as mudanças no estado cerebral, com o impacto afetado por propriedades específicas dos ramos, recrutamento diferencial de conjuntos pós-sinápticos e padrões de sincronização cerebrais inteiros. Isto se baseia em discussões de longo prazo sobre topologias de ramificação axonal e como estas poderiam modular o processamento de informação por atrasos de tempo na propagação de impulsos, filtragem diferencial específica de ramos e excitabilidade dependente de atividade (por exemplo, Segev e Schneidman, 1999). Com poucas exceções, como a via do tronco cerebral auditivo, ainda faltam dados concretos sobre ativações síncronas e assíncronas através de ramos filhos e como essas relações temporais podem impactar na resposta neuronal pós-sináptica (mas, veja Gomez-Laberge et al., 2016; Stoelzel et al., 2017). Assim, um desafio contínuo é elucidar características específicas dos ramos dentro dos axônios individuais e os efeitos sobre os conjuntos pós-sinápticos. Trabalho recente traz à tona questões adicionais sobre heterogeneidade da rede, incluindo por que os neurônios de uma única área de origem projetam variavelmente para um ou mais alvos no que está sendo descrito repetidamente como “em todas as combinações”
Contribuições do Autor
O autor confirma ser o único contribuinte deste trabalho e o aprovou para publicação.
Fundamento
Agradecemos aos Institutos Nacionais de Saúde (NIH) pelo apoio financeiro: MH107456.
Conflict of Interest Statement
O autor declara que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que pudessem ser interpretadas como um potencial conflito de interesses.
Barry, J. M. (2015). Atividade Axonal in vivo: considerações técnicas e implicações para a exploração de circuitos neurais em animais em livre movimentação. Frente. Neurosci. 9:153 doi: 10.3389/fnins.2015.00153
PubMed Abstract | CrossRef Full Text | Google Scholar
Gagnon, D., and Parent, M. (2014). Distribuição e VGLUT3 em eixos altamente colateralizados a partir do núcleo do ráfio dorsal do rato, conforme revelado por reconstruções de um neurônio. PLoS ONE 9:e87709. doi: 10.1371/journal.pone.0087709
CrossRef Full Text | Google Scholar
Kita, T., and Kita, H. (2012). O núcleo subthalâmico é um dos múltiplos locais de inervação para os axônios corticofugal de longo alcance: um único estudo de traçado de axônios no rato. J. Neurosci. 32, 5990-5999 doi: 10.1523/JNEUROSCI.5717-11
PubMed Abstract | CrossRef Full Text | Google Scholar