Frontiers in Systems Neuroscience
Förändringar i hjärnans tillstånd är förknippade med förändringar i mikrokretsar i neuronala brinnande egenskaper och med förändringar på makronivå i synkrona eller asynkrona mönster av hjärnans aktivering. Inom denna ram har axoner varit relativt mindre undersökta (Barry, 2015), och då främst i samband med ledningshastighet för aktionspotentialer och den ökade vakenhet som kan uppstå till följd av förändringar i ledningshastighet (t.ex. Stoelzel et al., 2017). Dessutom verkar dock den starkt förgrenade topologin hos många axoner vara väl utformad för att differentiellt rekrytera distribuerade postsynaptiska neuronala grupper, möjligen på ett tillståndsberoende sätt. Signalbehandling kan ske i minst tre axonala domäner; nämligen kartläggning, förstärkning och timing (Innocenti et al., 2016; Innocenti, 2017). I det här perspektivet diskuterar jag kortfattat först nätverksexempel på axonkollateralisering och sedan några av de axonintrinsiska egenskaper som kan ligga till grund för differentiell postsynaptisk rekrytering. I avsaknad av detaljerade parametrar eller mekanismer är målet främst att belysa generella egenskaper som kan figurera i kontroll och övergångar av hjärnans tillstånd.
Axonkollateraler
Alla axoner har en utstuderad distal arborisering i målstrukturen. Många axoner har dessutom flera grenar (även kallade collateraler) som riktar sig mot distinkt olika strukturer (granskad i Rockland, 2013, 2018). Ett utmärkt exempel är förgreningen av axoner från kortikothalamiska neuroner i lager 5. Detta har dokumenterats upprepade gånger och diskuteras ofta som en nyckelmekanism genom vilken en organism kan särskilja om förändringar i sensorisk input produceras av förändringar i miljön eller av självinitierade rörelser (”efference copy” eller ”collateral discharge”, Guillery och Sherman, 2011).
Inom gnagare, där det finns ett stort antal tillämpbara tekniker för att undersöka den här frågan, är kollateralisering känd för att vara relativt vanlig. För att ge några exempel: (1) Neuroner i hippocampus subiculum projicerar till mammillärkropparna, den retrospleniala cortexen eller genom kollateraler till båda (råtta: Kinnavane et al, 2018); (2) serotonerga neuroner i den dorsala raphekärnan (DR) kontaktar i olika kombinationer striatum, prefrontal cortex och amygdala (råtta: Gagnon och Parent, 2014) samt kombinationer av autonoma kärnor (Waselus et al., 2011). I det här fallet kan aktivering av stressresponsen genom DR-kollateraler uppnå synkroniserad aktivering av kärnor som är förknippade med neurohormonfrisättning eller pressoreaktioner. Differentiella, samordnade aktiveringar av strukturer i framhjärnan kan bidra till de mångfacetterade men relaterade DR-funktionerna, t.ex. reglering av sömn-väckningscykeln, modulering av smärtsignaler eller humöruttryck (Gagnon och Parent, 2014).
För det tredje identifierar anatomiska studier av thalamokortikala projektioner hos möss ”multispecifika axoner” som förgrenar sig vitt och brett till begränsade domäner i separata kortikala (och subkortikala) områden. Dessa har antagits orkestrera den snabba uppkomsten och rekonfigurationen av rumsligt distribuerade, synkroniserbara neurala sammansättningar (Clasca et al., 2016).
Fjärde, en nyligen genomförd studie av kortikokortikala anslutningar med hjälp av axonspårning i hela hjärnan i musens visuella cortex fann att 23 av 30 neuroner kontaktade från två till sju andra kortikala områden. Som bekräftelse på detta resultat fann höggenomsnitts-DNA-sekvensering av genetiskt streckkodade neuroner att 44 % av 533 neuroner var multipelt projicerande (”broadcast neurons”, Han et al., 2018). Han et al. urskilde preliminärt två breda typer av projicerande neuroner, en mindre ”dedikerad” (uni-target) subpopulation, som samexisterar med en prevalens av ”broadcasting” (multipelt projicerande) celler. Skulle denna arkitektur kunna tjäna modulationer i kognitivt tillstånd och sensorisk bearbetning?
Non-stereotypt kollateralisering
Som påpekats ovan gör neuroner som kollateraliserar det i ett icke-stereotypt mönster. Inom en bestämd projektion (definierad av ursprunget) skickar neuronerna grenar till en diversifierad undergrupp av målområden (”i alla kombinationer”). Denna observation är så konsekvent att den kan betraktas som en regel, även om de detaljerade parametrarna inte har tabellerats. Den funktionella betydelsen är också oklar, men nya resultat tyder på att de heterogena och varierande ingångarna till en kortikal neuron (och i förlängningen kan vi härleda det till neurala sammansättningar) är viktiga när det gäller att driva variabilitet och förändringar i spiktågen över olika experimentella försök (Gomez-Laberge et al, 2016 och se nedan).
Intrinsic Collaterals
I alla arter har kortikala projektionsneuroner på långa avstånd, förutom enstaka eller flera extrinsiska mål, typiskt sett en utarbetad intrinsisk arborisering. Vissa kortikala neuroner har endast en intrinsisk (lokal) kollateralisering och vissa endast en extrinsisk (råtta: Kita och Kita, 2012). Även om det bara finns få uppgifter om de faktiska proportionerna av intrinsikala och extrinsikala arboriseringar (Parent et al., 2000; Rockland, 2018), tyder allt på att detta kommer att vara mycket varierande. Till och med inom systemet med intrinsiska förbindelser finns det inte bara skillnader i antalet kollateraler och antalet synaptiska boutoner, utan en enskild neuron kan ha en blandning av myeliniserade och icke-myeliniserade kollateraler (kattens visuella cortex: Martin et al., 2014; Koestinger et al., 2017). Grenspecifik myelinisering skulle kunna resultera i ökad, grenspecifik ledningshastighet, även om Koestinger et al. antyder att det kan ha mer att göra med faktorer som ökad säkerhet i överföringen, förmodligen återigen grenspecifik.
En märklig observation relaterad till myelinisering gäller Gennaris stria, det myeliniserade bandet av axoner i skikt 4B i primaternas område V1. Eftersom detta består av inneboende kollateraler är den vanliga förklaringen, att myelinisering är ett sätt att öka ledningshastigheten över långa avstånd, inte omedelbart tillämplig. Lokala kollateraler behöver inte (?) ökas i förhållande till avlägsna mål, och i själva verket verkar det paradoxalt med en ökad lokal ledning (via myelinisering?) i förhållande till den synkronicitet som man hade kunnat förutsäga. Kan det vara så att myeliniseringen relaterar till andra faktorer, t.ex. plasticitetsrelaterade (eller tillståndsrelaterade) förändringar i axondiametern?
Ett intressant exempel: Meynert-celler i område V1 hos icke-mänskliga primater
Dessa stora neuroner, vid gränsen mellan lager 5 och 6, projicerar på varierande sätt till extrastriat område MT och/eller andra visuella associationsområden och/eller till pulvinarkärnan och/eller till superior colliculus och pretectum (Weisenhorn et al., 1995; Rockland och Knutson, 2001). Den inneboende kollateraliseringen är exceptionellt omfattande, uppmätt som 8,0 mm från soman, på grundval av subtotala rekonstruktioner, och med minst 800-1 370 boutoner (figur 1). Över hela den axonala arborisationen verkar det finnas en tydlig variabilitet i grendiameter; det vill säga, extrinsiska grenar som är riktade mot område MT är stora (diameter ~3,0 μm), men de intrinsiska grenarna och de som projicerar till pulvinar och superior colliculus verkar mindre, vilket bedöms med ljusmikroskopi (jfr. figur 1B (intrinsisk) och figur 1D (extrinsisk till område MT). Varierande diametrar tyder förmodligen på skillnader i graden av myelinisering och, som en följd därav, i ledningshastighet.
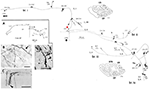
Figur 1. En typisk, rumsligt utsträckt proximal axonal arborisering av en Meynertcell (röd asterisk) i primär visuell cortex hos en makakapa. Det finns tre större intrinsiska kollateraler (märkta som br. i, ii, iv) som sträcker sig 3,0 mm dorsalt i lager 1, 3,0 mm ventralt i lager 6, 4,0 mm ventralt i lager 6 och 0,5 mm i lager 4B, vilket indikeras av pilspetsar i de två koronala sektionernas konturer (sektionerna 222 och 274, där dorsalt är till vänster). Sammanlagt sträcker sig de inneboende kollateralerna över 5,9 mm anterior-posterior (117 sektioner × 50 μm). Ytterligare en extrinsisk kollateral (br. iii) förekommer i lager 4 och 6 i område V2. Delar av de enskilda kollateralerna och av huvudaxonet (tjock pil) kunde inte följas, vilket anges med streckade linjer. Insatsen med låg förstoring (A, till vänster) ger en schematisk översikt av den allmänna konfigurationen. Siffrorna anger enskilda sektioner, där 20 siffror = 1,0 mm. Alla grenar har många små synaptiska kluster, varav ett illustreras i (B). Observera minskad diameter mellan huvudaxonet (pilen) och den terminala arboriseringen. Extrinsiska axoner (C) har varierande diameter (en axon med stor diameter i pilspetsen). (D) Extrinsiska termineringar i område MT omfattar några axoner med stor diameter. Skalstreck = 25 μm i (B), 100 μm i (C), 20 μm i (D). CF, calcarine fissure; LS, lunate sulcus; STS, superior temporal sulcus; L, layer. Modifierat från figurerna 1, 9 i Rockland och Knutson (2001) och figurerna 6f, 7b från Rockland (1995) med tillstånd.
Funktionell betydelse av förgrening av axon
En viktig komponent av kollateralisering är att dottergrenarna ofta inte är enhetliga, utan framför allt vid förgreningspunkterna varierar i diameter (figur 2). Variationen i diameter tillsammans med andra parametrar kommer att påverka excitabilitet, ledningshastighet och andra aspekter av signalspridning. Andra parametrar som påverkar är myelinets bredd och intermodal längd samt täthet och fördelning av jonkanaler (granskad i Debanne et al., 2011; Seidl, 2014; Bucher, 2016; Seidl och Rubel, 2016; Rama et al., 2018). Dessa skulle ha effekter på neurala responsegenskaper på mikrokretsnivå. På den mer globala nivån kan summerad aktivitet från flera projicerande axoner, med varierande ledningshastigheter över ett sammanlänkat nätverk, resultera i ett spektrum av synkrona och/eller asynkrona aktiveringar (Mitra et al., 2015; Zeki, 2016). Differentiell rekrytering av postsynaptiska populationer eller nätverksrekombinationer skulle kunna vara faktorer vid tillståndsövergångar eller modulering.
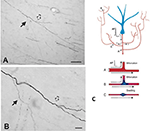
Figur 2. Ett typiskt förgrenat segment av axon i den vita substansen (makakapa). Segmentet har sitt ursprung från en neuron i parietal cortex och ses här i närheten av ventral temporal cortex. Panel (A) är en lägre förstoring av (B). Observera dubbla bifurkationer, där den första dottergrenen (heldragen pil) är påfallande tunnare (och omyeliniserad?) än huvudaxonet. I den andra, något mer distala bifurkationen (ihålig pil) verkar dottergrenarna vara ungefär lika stora i diameter, men båda är tunnare än huvudaxonet. Skalstreck = 100 μm (A) och 10 μm (B). Återgivna från Zhong och Rockland (2003) med tillstånd. (C) Schematisk bild av en neuron (blå) och dess förlängda förgreningstopologi (förkortat för att underlätta formateringen). En aktionspotential (AP) kan följa kretsvägar (A-C) till flera mål. Utbredningens tillförlitlighet beror både på axonets aktiva elektriska egenskaper och dess geometri, inklusive membraninhomogeniteter som svullnader och inkompatibla grendiametrar. Nedan: Schematisk bild för att illustrera tillförlitlig utbredning (A, med optimal impedansmatchning mellan moder- och dottergrenarna) och långsammare eller misslyckad utbredning (B, där dottergrenen har en större diameter; C, där det finns en mellanliggande membranuppsvällning). Återgivet från Huguenard (2000) med tillstånd.
Axonets geometri, aktiva elektriska egenskaper och membraninhomogeniteter vid grenarna är välkända som faktorer för tillförlitlig utbredning (t.ex. Manor et al., 1991; Innocenti et al., 1994; Tettoni et al., 1998; Huguenard, 2000; Ofer et al., 2017). Detta leder till flera olika scenarier om tidsmässiga egenskaper till följd av kollateralisering.
För det första kan det förekomma synkron aktivering i alla dottergrenar. De auditiva hjärnstamsbanorna projicerar via ett enda bifurkationsaxon till ipsi- och contralaterala mål (respektive korta och längre fysiska banor). Isokronisk överföring uppnås genom differentiell myelinisering och axonkaliber i de två dottergrenarna (dvs. mindre kaliber och kortare intermodala längder ipsilateralt; Seidl, 2014; Seidl och Rubel, 2016). Eftersom de flesta förgrenade axon, i jämförelse med hjärnstammens hörselbanor, täcker ett större område och understödjer mindre väldefinierade funktioner, är data till stor del ofullständiga eller saknas för andra system. Man skulle dock kunna förutsäga, när det gäller synkronicitet, att proximala grenar (dvs. de intrinsikala kollateralerna hos Meynert eller andra pyramidala celler) skulle ha anatomiska specialiseringar som resulterar i längre ledningstider, för att kompensera för och matcha de längre avstånden hos de extrinsikala kollateralerna. Såsom påpekats ovan tycks denna enkla förutsägelse inte hålla. Ytterligare undersökningar kommer att innebära provtagning från identifierade axoner över långa avstånd, vilket inte skulle vara lätt att uppnå.
För det andra kan grenspecifik aktivering vara asynkron. Detta kan bero på selektivt misslyckande i överföringen och/eller asynkrona ledningstider över axonets arbor (figur 2; Huguenard, 2000; Bucher, 2016). Modeller av kortikala kretsar beskriver distinkta routingtillstånd av kortvarig övergående synkronitet som dynamiskt skulle kunna forma informationsflödet (Palmigiano et al., 2017). Jämförbara experimentella data saknas i stort sett för axoner på långa avstånd. Den kollaterala topologin hos thalamokortikala projektioner ger dock ett exempel som framkallar ett komplicerat, aktivitets- och/eller tillståndsberoende asynkront aktiveringsmönster.
Kortikal och thalamokortikal aktivitet är i hög grad tillståndsberoende; och samspelet mellan presynaptiska extrinsiska inflöden (grenspecifika?) med inneboende membran- och synaptiska egenskaper hos postsynaptiska neuroner anses vara grundläggande för generering av rytmisk aktivitet (med ”vitt skilda effekter från förstärkning eller blockering av sensorisk-motorisk bearbetning….”, McCormick et al, 2015).
Variabilitet i kortikala svar är paradoxalt eftersom dessa också fungerar som substrat för stabila sensoriska upplevelser. Neuronal variabilitet har förknippats med graden av heterogen synkronisering över extrinsisk input; det vill säga, sensorisk stimulering eller beteendeförhållanden som ökar inputhomogeniteten till ett visst område förutspås också minska den neuronala variabiliteten (Gomez-Laberge et al., 2016). Nya diskussioner om mikrokretsresponser har spekulerat om en framträdande roll för små variationer eller skillnader i information: ”Men vad händer om skillnaderna mellan konnektiviteten inom kohorter av celler av samma klass är viktiga för kretsens funktion?”. (Morgan och Lichtman, 2017).
Dynamiska axonegenskaper
Förändringar i svarslatens har rapporterats i samband med olika tillstånd av vakenhet. I den kortikotalamiska banan leder ökad vakenhet till betydligt kortare svarstid. Detta och/eller förändringar i eldningsfrekvensen för ankommande impulser kan vara ansvariga för en dramatiskt ökad svarstillförlitlighet för subpopulationen (58 %) av visuellt responsiva kortikogeniculära neuroner (i kaniner: Stoelzel et al., 2017). Dessa resultat gäller fysiologiskt identifierade enskilda axoner; men man kan spekulera om en bredare tillämplighet på grenar av kollateraliserade axoner.
Löpande processer av synaptogenes och distal axonomsättning har påvisats i den vuxna hjärnbarken (NHP: Stettler et al., 2006). På kortare tidsskalor visar superupplösningsmikroskopi av icke-myeliniserad GFP-märkt CA3-hippocampus i organotypiska hjärnskivor att axonerna gradvis vidgas efter anfall av högfrekvent eldning, en observation som bekräftas av elektrofysiologisk registrering (Chereau et al., 2017). Andra, grenspecifika förändringar kommer sannolikt att upptäckas; till exempel har terminala arboriseringar av individuellt märkta axoner från dorsal raphe en målspecifik andel boutoner som innehåller proteinet VGLUT3 (större andel för grenar som avslutas i striatum än i motorisk cortex). Detta innebär en komplex, icke enhetlig trafikeringsmekanism över collateraler (Gagnon och Parent, 2014).
Slutsats
I denna Perspective-artikel har jag diskuterat axonförgrening som relevant för förändringar i hjärnans tillstånd, med påverkan som sker via grenspecifika egenskaper, differentiell rekrytering av postsynaptiska ensembler och hela hjärnans mönster av synkronisering. Detta bygger på långvariga diskussioner om axonala förgreningars topologier och hur dessa kan modulera informationsbearbetning genom tidsfördröjningar i impulsutbredning, differentiell grenspecifik filtrering och aktivitetsberoende excitabilitet (t.ex. Segev och Schneidman, 1999). Med bara några få undantag, t.ex. den auditiva hjärnstamsbanan, saknas fortfarande i stort sett hårda data om synkrona och asynkrona aktiveringar genom dottergrenar och hur dessa tidsmässiga förhållanden kan påverka postsynaptisk neuronal responsivitet (men, se Gomez-Laberge et al., 2016; Stoelzel et al., 2017). En fortsatt utmaning är således att belysa grenspecifika egenskaper inom enskilda axoner och effekterna på postsynaptiska ensembler. Nya arbeten aktualiserar ytterligare frågor om nätverksheterogenitet, inklusive varför neuroner från ett enda källområde varierar i projektion till ett eller flera mål i vad som upprepade gånger beskrivs som ”i alla kombinationer.”
Författarbidrag
Författaren bekräftar att han eller hon är den enda bidragsgivaren till detta arbete och har godkänt det för publicering.”
Finansiering
Vi tackar National Institutes of Health (NIH) för finansiellt stöd: MH107456.
Intressekonfliktförklaring
Författaren förklarar att forskningen utfördes i avsaknad av kommersiella eller finansiella relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Barry, J. M. (2015). Axonal aktivitet in vivo: tekniska överväganden och konsekvenser för utforskandet av neurala kretsar hos fritt rörliga djur. Front. Neurosci. 9:153 doi: 10.3389/fnins.2015.00153
PubMed Abstract | CrossRef Full Text | Google Scholar
Gagnon, D. och Parent, M. (2014). Distribution og VGLUT3 i högt kollateraliserade axoner från råttans dorsala raphe-kärna som avslöjas av rekonstruktioner av enskilda neuroner. PLoS ONE 9:e87709. doi: 10.1371/journal.pone.0087709
CrossRef Full Text | Google Scholar
Kita, T. och Kita, H. (2012). Den subthalamiska kärnan är en av flera innervationsplatser för kortikofugala axoner med lång räckvidd: en studie av spårning av enstaka axoner hos råtta. J. Neurosci. 32, 5990-5999 doi: 10.1523/JNEUROSCI.5717-11
PubMed Abstract | CrossRef Full Text | Google Scholar