Clé vétérienne
- La famille des Anatidés, sous-famille des Anatinae, par exemple le canard colvert, le canard pelleteur, l’eider et les tadornes
- La famille des Anatidés, sous-famille des Anserinae, Tribu Anserini (cygnes et oies véritables), par exemple les cygnes tuberculés, chanteurs et de Bewick, les oies bernaches et les oies cendrées
- La famille des Falconidae par exemple le faucon pèlerin, le saker, le llanner, le faucon gerfaut
- La famille des Accipitridae comme les buses (commune, à pattes rudes et à miel), l’épervier, l’autour, l’aigle royal
Les Psittaciformes sont parmi les plus colorés des oiseaux gardés comme animaux de compagnie.
Système nerveux
Le cerveau aviaire est extrêmement lisse, dépourvu des nombreux gyri (les crêtes du cerveau) observés chez les mammifères (figure 9.1). La vue semble être le sens dominant chez la plupart des oiseaux. Deux grands lobes optiques se trouvent entre les hémisphères cérébraux et le cervelet, et c’est là que les nerfs optiques communiquent et diffusent les informations. Il n’y a pas de corps calleux, le cortex cérébral est généralement très fin, mais le corps strié est bien développé et on pense qu’il est le siège des associations mentales chez les oiseaux. Une caractéristique importante du cerveau de l’oiseau est le corps pinéal qui se trouve dans le diencéphale, cranial au cervelet, sur la ligne médiane dorsale. Le corps pinéal possède des cellules sécrétrices similaires aux photorécepteurs et réagit donc à la lumière. Il est également relié au nerf optique par les ganglions cervicaux crâniens. Le corps pinéal est responsable de la régulation de nombreux effets saisonniers tels que la reproduction et la migration, ainsi que des rythmes circadiens. Il a un effet direct via la sécrétion d’hormones sur l’hypothalamus.
Figure 9.1 Aspect dorsal des hémisphères cérébraux aviaires montrant l’absence de gyri.

Le système nerveux aviaire n’est pas différent de celui observé chez son homologue mammifère. Les oiseaux possèdent 12 nerfs crâniens (CN), soit le même nombre que chez le chat et le chien. Chez les oiseaux, le nerf optique (CN II) est le plus grand nerf crânien, avec un diamètre presque égal à la moitié de celui de la colonne vertébrale. Il traverse le calvarium par un seul trou plutôt que par de multiples petits trous associés à la plaque criblée des mammifères. Les 11 autres nerfs crâniens ont des fonctions similaires à celles observées chez les mammifères, et le lecteur est renvoyé à des textes d’anatomie plus approfondis pour plus d’informations (King &McLelland, 1975 ; Bennett, 1994).
Chacune des ailes est alimentée nerveusement par un plexus brachial dérivé des nerfs spinaux dans la zone cervicale caudale. Il existe trois plexus nerveux dans la région lombosacrée : lombaire, ischiatique et pudendal. Le plexus lombaire dérive des deux dernières racines nerveuses lombaires et de la première ou des deux premières racines nerveuses sacrées. Comme les autres plexus lombosacrés, il se situe dans un creux du bassin, en position dorsale par rapport à la région rénale crânienne. Il alimente les muscles de la paroi corporelle et de la partie supérieure de la jambe et donne naissance aux nerfs obturateur, fémoral, fessier crânial et saphène. Contrairement aux chiens et aux chats, les oiseaux possèdent un plexus ischiatique qui dérive de quatre à sept nerfs spinaux dans la région sacrée et qui est situé dans un creux du bassin, dorsalement à la structure rénale moyenne. Il donne naissance aux principales sources nerveuses des membres postérieurs – le nerf ischiatique, qui est le plus grand nerf périphérique du corps, et le nerf fessier caudal. Enfin, un plexus pudendal se forme dans un creux du bassin dorsal à la région rénale caudale à partir de cinq nerfs spinaux coccygiens et innerve la queue et la région cloacale.
Système musculo-squelettique
La plupart des oiseaux ont le pouvoir de voler. Les os denses et encombrants du mammifère terrestre demanderaient trop d’efforts pour être soulevés dans les airs. Les oiseaux ont donc adapté leur squelette, simplifiant le nombre d’os en fusionnant certains, et allégeant généralement l’ensemble de la structure en créant des espaces d’air à l’intérieur de nombreux os.
Pour alléger encore le squelette, plusieurs des plus gros os, et même certaines des vertèbres de la colonne vertébrale, sont reliés directement ou indirectement aux voies respiratoires, et sont dits pneumonisés. Le tissu osseux, qui remplace l’épaisse cavité médullaire ou moelle osseuse présente au centre des os des mammifères, présente une structure légère et trabéculaire. Bien que légère, cette structure est néanmoins extrêmement solide.
La figure 9.2 montre un squelette aviaire généralisé.
Figure 9.2 Squelette aviaire.
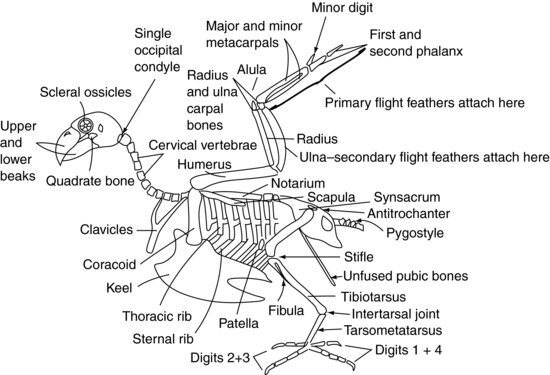
Crâne
Bec
Le bec, ou bill, est la principale caractéristique du crâne aviaire. Il a été modifié en un nombre ahurissant de formes et de tailles, dépendant principalement du régime alimentaire auquel l’oiseau s’est adapté. Dans tous les cas, il est composé d’un bec supérieur (maxillaire) et d’un bec inférieur (mandibulaire) qui sont recouverts d’une couche de kératine, un composé protéique résistant similaire à celui qui forme l’exosquelette des insectes. Cette couche de kératine est connue sous le nom de rhamphothèque. Elle est ensuite classée de manière à ce que la couche maxillaire soit appelée rhinothèque, et la couche mandibulaire gnatothèque. Le rhinothèque et le gnatothèque se développent à partir d’une plaque située à la base des côtés respectifs du bec, le taux de remplacement dépendant du type de nourriture consommée et de l’abrasion que reçoit le bec.
Chez les Psittaciformes (tableau 9.1), le bec supérieur est puissamment développé et se termine par une pointe acérée surplombant le bec inférieur plus large et plus robuste. L’énorme puissance du bec d’un perroquet est due à une articulation synoviale ou à un mécanisme de charnière, appelé articulation cinétique, qui relie le bec au crâne. Le bec inférieur du perroquet possède une série de capteurs de pression à son extrémité, qui lui permettent de tester la consistance et la structure des objets saisis.
Chez les rapaces, le bec supérieur est extrêmement pointu et tranchant, mais il est dépourvu de l’attache de l’articulation cinétique et ne peut donc pas produire une force descendante aussi puissante. Au lieu de cela, il est utilisé comme un instrument de déchirure.
Chez les Ansériformes (la famille des canards), le bec est aplati et peut avoir de fines dentelures sur les bords qui permettent à l’oiseau de filtrer les fines particules de l’eau. Les canards tels que les canards colverts et les canards de surface ont ce type de bec. Ces dentelures peuvent être développées jusqu’à un bord dentelé (par exemple, dans la famille bien nommée des becs-scie), ce qui permet à l’oiseau de saisir des aliments glissants, comme le poisson. Les Ansériformes possèdent également des terminaisons nerveuses dans une plaque située à l’extrémité de leur bec (appelée » ongle « ) qui leur permet de trouver de la nourriture cachée dans la boue.
Chez tous les oiseaux, il existe une série de petits os derrière les becs inférieur et supérieur qui leur permettent de déplacer le bec indépendamment du crâne. Il s’agit notamment des os palatin, quadrate et ptérygoïde et des arcs jugaux. Leurs mouvements exacts dépassent le cadre de ce texte pour être décrits, mais de nombreuses références à la fin de ce chapitre donnent de bons comptes rendus de leur fonction.
Nostrils
Les narines, ou narines, se trouvent à la base du bec supérieur chez la plupart des oiseaux et sont souvent entourées d’une zone de peau sans plumes appelée la céré. Celle-ci peut être très colorée chez certaines espèces, comme la perruche, où elles peuvent être utilisées pour identifier le sexe de l’oiseau. Chez de nombreux Anseriformes, les narines sont situées plus près de l’extrémité du bec. Les narines elles-mêmes ne sont que des ouvertures vers les chambres sinusales, qui sont elles-mêmes reliées à un réseau ramifié de chambres osseuses dans toute la tête de l’oiseau. Ces sinus varient selon l’espèce, mais la majorité des patients aviaires ont un sinus infra-orbitaire. Celui-ci se situe sous les yeux et est souvent impliqué dans les infections sinusales et oculaires. Il se distingue des sinus observés chez la plupart des mammifères par l’absence d’os dans la paroi latérale, qui n’est recouverte que de tissus mous. Cela signifie que toute infection du sinus infra-orbitaire se traduit souvent par un gonflement sur la face de l’oiseau, en position ventrale par rapport à l’œil. Ces sinus communiquent également avec les sacs aériens de la tête et du cou. La fonction de ces sacs aériens n’est pas claire, mais ils peuvent aider à la résonance de la voix. Lorsqu’un oiseau souffre d’une infection des sinus, les entrées étroites de ces sinus peuvent être partiellement bloquées et agir comme des valves à sens unique, permettant à l’air d’entrer dans les sacs mais pas d’en sortir. Les sacs peuvent alors se gonfler de façon excessive et des gonflements mous sont alors fréquemment observés sur l’arrière ou la nuque de l’oiseau.
Les sinus et les narines externes communiquent avec l’oropharynx par la fente choanale. Il s’agit d’une ouverture étroite sur la ligne médiane du palais dur, située immédiatement au-dessus de la glotte lorsque le bec est fermé, permettant à l’oiseau de respirer par les narines. C’est souvent la zone choisie pour prélever des échantillons lorsqu’on essaie d’isoler des agents infectieux pour les maladies des voies aériennes supérieures chez les oiseaux.
Le crâne du patient aviaire se connecte à l’atlas (ou première vertèbre spinale) via un seul condyle occipital à la base du crâne, contrairement aux deux des mammifères. Il existe également un grand nombre de vertèbres cervicales très mobiles. Ces deux facteurs rendent la tête aviaire extrêmement agile. Cependant, l’articulation atlanto-occipitale est également un point faible, ce qui rend la luxation à cet endroit très facile.
Colonne vertébrale
Vertébres cervicales
Les vertèbres cervicales (figure 9.2) sont mobiles de façon indépendante chez l’aviaire, comme chez le mammifère, et leur nombre varie selon les espèces entre 11 et 25. Elles ont généralement une forme de boîte.
Vertébres thoraciques, lombaires et sacrées
Les vertèbres thoraciques (figure 9.2) sont fusionnées chez les rapaces, les pigeons et de nombreuses autres espèces pour former un seul os appelé notarium. Chez d’autres espèces, elles ont une mobilité limitée. Il y a ensuite deux articulations intervertébrales entre le notarium et les vertèbres lombaires et sacrées fusionnées. Ces vertèbres soudées sont appelées synsacrum. Le synsacrum fusionne avec le bassin lui-même pour former un bouclier osseux dorsal sur la face caudale de l’oiseau.
Vertébres coccygiennes
La majorité des vertèbres coccygiennes caudales (figure 9.2) sont généralement fusionnées en une seule structure connue sous le nom de pygostyle – qui forme la partie « nez de perse » du poulet!
Pelvis
Le toit du pelvis est formé par le synsacrum (figure 9.2). Les deux » côtés » du bassin sont de taille réduite par rapport aux mammifères mais sont constitués des os iliaques et ischiatiques, l’acétabulum étant créé à leur point de rencontre. Chez les oiseaux, l’acétabulum n’est pas une cavité osseuse complète comme chez les mammifères, mais une feuille fibreuse. Il existe une crête sur le bassin latéral, appelée antitrochanter, qui s’articule avec le grand trochanter du fémur. La fonction de cette crête est d’empêcher l’abduction du membre lors du perchage. Les os pubiens du bassin ne fusionnent pas sur la ligne médiane ventrale comme chez les mammifères. Ils forment plutôt de fins os longs qui s’étendent caudalement vers l’évent. Ils fournissent un support à la peau recouvrant l’abdomen caudal et un espace suffisant pour le passage des œufs chez l’oiseau femelle.
Cage thoracique
Les pittaciformes possèdent huit paires de côtes (figure 9.2). Chaque côte a un segment dorsal appelé côte thoracique, et un segment ventral, ou côte sternale. Ces côtes pointent vers l’arrière et relient rigidement les vertèbres thoraciques dorsalement et la quille, ou sternum, ventralement.
Sternum
Les vertèbres sternales sont fusionnées chez les oiseaux pour former la quille. La quille présente une crête médiane qui divise les muscles pectoraux en côtés droit et gauche. La crête peut être une structure profonde, comme on le voit chez les pigeons, les rapaces et les Psittaciformes, permettant aux grands muscles pectoraux de se fixer pour un vol puissant. Alternativement, la quille peut être aplatie, comme chez les Anseriformes, afin de fournir une structure semblable à celle d’un bateau, plus adaptée à la flottaison.
Ailes
L’articulation de l’épaule est formée par la réunion de trois os : l’humérus, l’omoplate (qui est plus tubulaire que celle aplatie des mammifères) et un troisième os appelé coracoïde (figure 9.2). Ce dernier os forme une entretoise qui soutient l’articulation de l’épaule contre le sternum. Le muscle supracoracoïde s’attache à la quille, puis passe par le foramen, ou ouverture, formé au point de rencontre de ces os, et atteint ainsi la face dorsale de l’humérus où il s’attache. La contraction de ce muscle, ainsi que de certains tissus élastiques également présents, permet de soulever l’aile. Les muscles pectoraux s’attachent de la quille à l’humérus pour tirer l’aile vers le bas. Les clavicules soudées, ou bréchet (souvent appelé furcula), s’articulent avec l’os coracoïde et confèrent un certain ressort au battement des ailes. L’humérus est pneumonisé, ce qui signifie qu’il ne peut pas être utilisé pour la fluidothérapie intra-osseuse. C’est également un point important à prendre en compte lors de la réparation de fractures.
L’humérus s’articule avec le radius et le cubitus au niveau de l’articulation du coude. Le radius est le plus petit de ces deux os et se situe crânialement. Le cubitus constitue la source d’attache des plumes de vol secondaires, qui s’insèrent directement dans le périoste de cet os (figure 9.3). Le cubitus est souvent utilisé pour l’administration intra-osseuse de fluides chez les oiseaux.
Figure 9.3 Aspect ventral de l’aile d’une crécerelle (Falco tinnunculus) avec les plumes de couverture enlevées montrant l’attachement des primaires au manus et des secondaires au cubitus.

Le radius et le cubitus s’articulent respectivement avec un os carpien radial et un os carpien ulnaire. Ceux-ci s’articulent à leur tour avec trois os métacarpiens. Le premier os métacarpien est l’équivalent du « pouce » aviaire. Il est connu sous le nom d’alula, ou « aile bâtarde », et forme une projection plumeuse à partir de la face crâniale de l’articulation carpométacarpienne. Les deux autres os métacarpiens sont connus sous le nom d’os métacarpiens majeurs et mineurs, et s’articulent avec la première phalange crânialement et le doigt mineur caudalement. La première phalange s’articule ensuite avec la deuxième phalange, formant ainsi l’extrémité de l’aile. Les plumes primaires s’attachent au périoste des phalanges et des métacarpiens mineurs (figure 9.3).
La surface de l’aile est agrandie par de fines feuilles de tissu élastique qui s’étendent d’une surface articulaire à l’autre. La plus grande s’étend de l’épaule à l’articulation du carpe crânialement et est connue sous le nom de propatagium ou » toile d’aile » (figure 9.4). Elle peut être utilisée chez certaines espèces, comme les pigeons, pour l’administration de vaccins.
Figure 9.4 Aspect dorsal de l’aile d’une crécerelle (Falco tinnunculus) avec les plumes de couverture enlevées montrant la feuille élastique du propatagium pontant l’articulation du coude.

Membre pelvien
L’acétabulum du pelvis maintient la tête fémorale (figure 9.2). Le membre peut être verrouillé, et empêché d’être abducté, par l’engagement du grand trochanter du fémur avec la crête antitrochantérienne du bassin. Le fémur est pneumonisé chez de nombreux oiseaux. Au niveau de l’articulation du grasset, le fémur s’articule avec la rotule et l’os tibiotarsal. L’os tibiotarsal est appelé ainsi parce qu’il est formé par la fusion du tibia et de la rangée proximale des os du tarse, et peut également être utilisé pour l’administration intra-osseuse de fluides. Sur la face latérale du tibiotarse proximal se trouve la fibula, très réduite.
Distalement, l’os tibiotarse s’articule avec l’os tarsométatarsien. Cet os est formé par la fusion de la rangée distale des os du tarse avec l’os métatarsien solitaire. L’articulation entre le tibiotarse et le tarsométatarse est appelée articulation intertarsale, ou suffragette. Le tarsométatarse s’articule ensuite avec les phalanges.
Chez les Psittaciformes, deux doigts pointent vers l’avant (le deuxième et le troisième) et deux vers l’arrière (le premier et le quatrième), créant un membre zygodactyle. Le premier doigt a deux phalanges, le deuxième doigt a trois phalanges, le troisième a quatre phalanges et le quatrième a cinq phalanges. Chez les oiseaux percheurs (Passeriformes) et les rapaces, les deuxième, troisième et quatrième doigts pointent vers l’avant et le premier vers l’arrière, créant ainsi un membre anisodactyle. Certaines espèces, comme le balbuzard pêcheur (Pandion haliaetus), peuvent déplacer le quatrième doigt pour qu’il soit orienté vers l’avant ou vers l’arrière afin de faciliter la capture de sa proie, créant ainsi un membre semi-zygodactyle.
Sens particuliers
Oeil
L’œil aviaire est unique car il contient une série de petits os. Ceux-ci sont connus sous le nom d’osselets scléraux (figure 9.2). Ils forment une structure en forme d’anneau qui soutient l’avant de l’œil. L’œil aviaire diffère également de l’œil des mammifères en ce qu’il n’est pas un globe, mais en forme de poire, avec l’extrémité la plus étroite à l’extérieur.
L’œil aviaire est grand par rapport à la taille globale du crâne, avec seulement un septum osseux mince comme du papier qui sépare les orbites droite et gauche. Les oiseaux possèdent une troisième paupière mobile et translucide, ainsi que des paupières supérieures et inférieures, l’inférieure étant plus mobile que la supérieure. Deux glandes productrices de larmes existent couramment : la troisième paupière, ou glande de Harder, qui est située à la base de la troisième paupière, et la glande lacrymale située caudolatéralement, comme chez les mammifères.
La couleur de l’iris peut changer avec l’âge chez certains perroquets, par exemple le perroquet gris d’Afrique a un iris gris foncé jusqu’à l’âge de 4-5 mois, où il devient jaune-gris, puis argenté en continuant à vieillir. Chez d’autres, l’iris peut servir d’indicateur du sexe de l’oiseau : chez les grands cacatoès, par exemple, la femelle a un iris rouge-brun brillant, alors que celui du mâle est brun-noir foncé.
La rétine aviaire est épaisse et ne possède pas de vaisseaux sanguins de surface visibles, contrairement à celle des mammifères. Pour assurer la nutrition de la rétine, les oiseaux possèdent une structure vasculaire plissée et repliée appelée le pecten oculi, qui se trouve à l’endroit où le nerf optique pénètre dans l’œil. Il se contracte par intermittence, expulsant les nutriments dans l’humeur vitrée.
Enfin, l’iris aviaire possède des fibres musculaires squelettiques en son sein, contrairement aux mammifères qui ne possèdent que des fibres musculaires lisses. Cela signifie que le patient aviaire peut contracter et dilater sa pupille à volonté, réduisant ainsi la valeur du réflexe lumineux pupillaire comme outil de détermination de la fonction oculaire. Comme les deux nerfs optiques sont complètement séparés l’un de l’autre, le réflexe lumineux consensuel est également un mauvais indicateur de la fonction cérébrale.
Oreille
Il n’y a pas de pavillon chez les oiseaux, bien que certaines espèces, comme les hiboux à longues et courtes oreilles, aient des plumes dans cette zone. Il existe un canal externe court et horizontal, recouvert de plumes, qui est situé caudolatéralement à l’orbite oculaire. La membrane du tympan est clairement visible. L’oreille moyenne est reliée à l’oropharynx par le canal d’Eustache. Les osselets auditifs des mammifères sont remplacés chez l’oiseau par un cartilage latéral extra columellaire et un os columellaire médial qui transmettent les ondes sonores à l’oreille interne.
L’oreille interne contient la cochlée et les canaux semi-circulaires, qui remplissent les mêmes fonctions que chez les mammifères.
Anatomie respiratoire
Système respiratoire supérieur
Les narines s’ouvrent sur les voies nasales, qui communiquent à leur tour avec la glotte du larynx par une ouverture médiane dans le palais dur qui forme le toit de la bouche caudale. Cette ouverture s’appelle la fente choanale. Le système sinusal et les sacs aériens cervico-céphaliques ont été mentionnés précédemment.
Larynx
Les oiseaux ont une structure laryngée réduite, dépourvue d’épiglotte, du cartilage thyroïdien et des plis vocaux observés chez les chats et les chiens. La structure principale est la glotte, qui protège l’entrée de la trachée. Des muscles externes tirent la glotte et la trachée vers l’avant afin qu’elles communiquent directement avec la fente choanale, permettant à l’oiseau de respirer par les narines. La glotte est maintenue fermée au repos, ne s’ouvrant que lors de l’inspiration et de l’expiration.
Trachée
La trachée des espèces aviaires diffère de la trachée des mammifères en ce que ses anneaux cartilagineux sont des cercles complets, en forme d’anneau de signet, s’emboîtant l’un sur l’autre, plutôt que les anneaux en forme de C de la trachée des mammifères. Chez les Psittaciformes et les rapaces diurnes, la forme de ces anneaux cartilagineux est légèrement aplatie en direction dorsoventrale, alors que chez la plupart des Passeriformes, ils sont ronds.
Dans certaines espèces, comme le cygne chanteur et la pintade, la trachée forme une série de boucles et d’enroulements au niveau de l’entrée thoracique. Chez d’autres espèces, comme l’émeu, la trachée est divisée en deux sur la ligne médiane ventrale, à trois quarts de la distance entre la tête et l’entrée thoracique. La muqueuse de la trachée se projette à travers cette fente pour former un sac trachéal. Cela améliore la résonance vocale. Chez les canards mâles, comme les colverts, il y a un gonflement dans la dernière partie de la trachée, souvent juste à l’intérieur du thorax, connu sous le nom de bulle trachéale.
Syrinx
Avant que la trachée ne se divise en deux bronches principales, il y a une structure connue sous le nom de syrinx (figure 9.6). C’est là que l’oiseau produit la majeure partie de sa voix. Il est composé d’une série de muscles et de deux membranes qui peuvent être mises en vibration, indépendamment de l’inspiration ou de l’expiration.
Système respiratoire inférieur
Poumons
Les poumons des espèces aviaires ont une structure rigide et ne se gonflent ou se dégonflent pas de manière significative. Ils sont de forme aplatie et fermement fixés à la face ventrale des vertèbres thoraciques et des côtes vertébrales. Il n’y a pas de diaphragme chez les oiseaux, et la cavité corporelle commune est appelée le cœlome.
Les bronches jumelées sont soutenues par des anneaux de cartilage en forme de C, contrairement à la trachée. Les bronches primaires alimentent chacun des deux poumons, et se divisent rapidement en bronches secondaires et tertiaires, ou parabronches. Quatre groupes principaux de bronches secondaires alimentent le poumon, mais leur rôle dans les échanges gazeux est minime. Les bronches tertiaires, en revanche, jouent un rôle dans les échanges gazeux, car leurs parois sont remplies de membranes capables d’échanger des gaz. Ces zones apparaissent comme de petites fosses, ou atriums, auxquelles sont reliés des tubes encore plus fins, appelés capillaires aériens. Ceux-ci s’entrecroisent les uns avec les autres pour former un maillage tridimensionnel entrelacé avec les lits capillaires sanguins. Ces capillaires aériens sont de taille variable, mais leur diamètre moyen est d’environ 3 à 5 mm. Ce diamètre extrêmement réduit produit des forces d’attraction très élevées entre leurs parois en présence de sécrétions liquides, ce qui entraîne un blocage rapide des surfaces respiratoires. Pour empêcher cela, il y a des cellules dans les parabronchi qui sécrètent du surfactant, pour s’assurer que les voies respiratoires restent ouvertes.
La structure pulmonaire peut être encore classée selon la direction du flux d’air en son sein en poumon néopulmonaire et poumon paléopulmonaire. Ces derniers seront mentionnés plus tard lors de l’examen de la physiologie respiratoire.
Sacs aériens
La dernière partie du système respiratoire inférieur aviaire est composée des sacs aériens (voir figure 9.5). Ce sont des sacs en forme de ballons qui agissent comme des soufflets, pompant l’air dans et hors des poumons aviaires rigides en réponse aux mouvements de la paroi corporelle et du sternum. Les parois des sacs aériens sont très fines et composées d’un épithélium pavimenteux simple qui recouvre une couche de tissu conjonctif élastique peu vascularisé.
Figure 9.5 Système de sacs aériens aviaires chez un canard : (a) voies nasales ; (b) sinus infra-orbitaire ; (c) sacs aériens cervico-céphaliques (simples) ; (d) sacs aériens claviculaires ; (e) sacs aériens thoraciques crâniens ; (f) sacs aériens thoraciques caudaux ; et (g) sacs aériens abdominaux. H, cœur ; L, foie ; Lu, poumons ; B, bulle syringale (canards mâles).
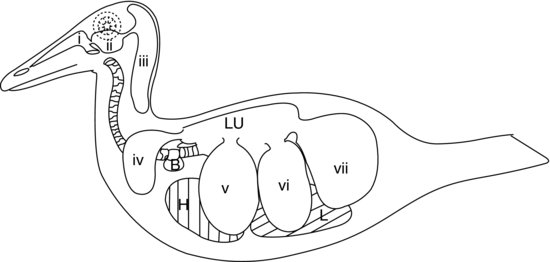
Dans la majorité des oiseaux, il existe neuf sacs aériens. L’un d’entre eux est le sac aérien séparé déjà mentionné, le sac aérien cervico-céphalique, qui ne communique pas du tout avec les poumons. Les huit autres communiquent tous avec les poumons via une bronche secondaire (à l’exception des sacs aériens abdominaux qui se connectent à la bronche primaire de chaque côté). La figure 9.5 montre le système de sacs aériens d’un canard.
En plus du sac aérien cervico-céphalique séparé, les huit autres sacs aériens standard sont
.