Chave Veterinária
- A família Anatidae, subfamília Anatinae, por exemplo o pato-real, pato pá, pato éider e pato shelducks
- A família Anatidae, subfamília Anserinae, Tribo Anserini (cisne e gansos verdadeiros), por exemplo o mudo, cisnes Whooper e Bewick, gansos barnacle e greylag
- A família Falconidae por exemplo o falcão peregrino, o padeiro, o lhanner, o gyrfalcon
- A família Accipitridae como os abutres (comuns, de pernas ásperas e mel), o pardal, o goshawk, o goshawk, a águia dourada
Os Psitacídeos estão entre os mais coloridos dos pássaros mantidos como animais de companhia.
Sistema nervoso
O cérebro aviário é extremamente liso, faltando os muitos gyri (as cristas do cérebro) vistos nos mamíferos (Figura 9.1). A visão parece ser o sentido dominante na maioria das aves. Dois grandes lóbulos ópticos encontram-se entre os hemisférios cerebrais e o cerebelo, e é aqui que os nervos ópticos comunicam e disseminam informações. Não há corpus callosum, o córtex cerebral é geralmente muito fino, mas o corpus striatum é bem desenvolvido e é considerado o local de associação mental nas aves. Uma característica importante do cérebro da ave é o corpo pineal que se encontra no diencéfalo, craniano até ao cerebelo na linha mediana dorsal. O corpo pineal possui células secretoras semelhantes às dos fotorreceptores e, portanto, responderá à luz. Elas estão ligadas também através dos gânglios cervicais cranianos ao nervo óptico. O corpo pineal é responsável por regular muitos efeitos sazonais como a reprodução e a migração, bem como os ritmos circadianos. Tem um efeito direto via secreção hormonal sobre o hipotálamo.
Figure 9.1 Aspecto dorsal dos hemisférios cerebrais aviários mostrando falta de gyri.

O sistema nervoso aviário não é diferente do observado em seu homólogo mamífero. As aves possuem 12 nervos cranianos (CN), o mesmo número que em cães e gatos. Nas aves, o nervo óptico (CN II) é o maior nervo craniano, sendo quase metade do diâmetro da coluna vertebral. Ele passa pelo calvário através de um único orifício e não através de vários orifícios menores associados à placa cribriforme dos mamíferos. Os outros 11 nervos cranianos têm funções semelhantes às observadas nos mamíferos, sendo o leitor encaminhado para textos de anatomia mais profunda para mais informações (King & McLelland, 1975; Bennett, 1994).
Cada uma das asas tem um suprimento nervoso de um plexo braquial derivado dos nervos espinhais na área cervical caudal. Existem três plexos nervosos na região lombossacral: lombar, isquiático e pudendo. O plexo lombar deriva dos dois últimos lombares e o primeiro a duas raízes do nervo espinhal sacral. Tal como os outros plexos lombossacrais, encontra-se numa cavidade da pélvis, dorsal até à zona do rim craniano. Fornece a parede do corpo e os músculos da coxa e dá origem aos nervos obturador, femoral, glúteo craniano e safena. Ao contrário dos cães e gatos, as aves têm um plexo isquiático que é derivado de quatro a sete nervos espinhais na área sacral e que está situado numa cavidade da pélvis dorsal até à estrutura média dos rins. Dá origem ao principal suprimento nervoso para os membros posteriores – o nervo isquiático, que é o maior nervo periférico do corpo e o nervo glúteo caudal. Finalmente, um plexo pudendo forma-se em uma cavidade da pelve dorsal até a área do rim caudal a partir de cinco nervos coccígeos espinhais e inerva a cauda e área cloacal.
Sistema músculo-esquelético
A maioria das aves tem o poder de voar. Os ossos densos e pesados dos mamíferos terrestres exigiriam muito esforço para se erguerem no ar. Assim, as aves adaptaram a sua estrutura esquelética, simplificando o número de ossos através da fusão de alguns, e geralmente clareando toda a estrutura criando espaços de ar dentro de muitos dos ossos.
Para clarear ainda mais o esqueleto, vários dos ossos maiores, e mesmo algumas das vértebras da coluna vertebral, estão ligados directa ou indirectamente às vias respiratórias, e diz-se que estão pneumonizados. Isto substitui a espessa cavidade medular ou medula óssea presente no centro dos ossos dos mamíferos, e produz uma estrutura leve e trabecular. Enquanto leve, a estrutura é, no entanto, extremamente forte.
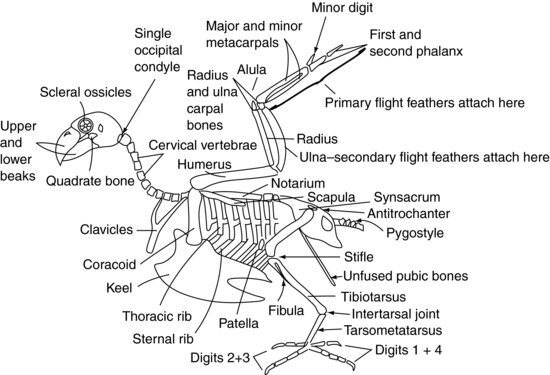
Figure 9.2 mostra um esqueleto aviário generalizado.
Figure 9.2 Esqueleto aviário.
Skull
Bico
O bico, ou bico, é a característica principal do crânio aviário. Foi modificado para um número desconcertante de formas e tamanhos, dependendo principalmente da dieta à qual a ave se adaptou. Em todos os casos é composto por um bico superior (maxilar) e inferior (mandibular) que são cobertos por uma camada de queratina, um composto proteico resistente semelhante ao que forma o exoesqueleto dos insectos. Esta camada de queratina é conhecida como rhamphotheca. É ainda classificada de forma a que a camada maxilar seja referida como rinoteca, e a camada mandibular como gnatotheca. A rinoteca e a gnatoteca crescem de um prato na base dos respectivos lados do bico, a taxa de reposição depende do tipo de alimento consumido e da abrasão que o bico recebe.
Em Psittaciformes (Tabela 9.1), o bico superior está poderosamente desenvolvido e termina em um ponto agudo sobre o bico inferior mais largo e mais gordo. O tremendo poder no bico de um papagaio deve-se a uma articulação sinovial ou mecanismo de dobradiça, conhecido como articulação cinética, que une o bico ao crânio. O bico inferior do papagaio tem uma série de sensores de pressão na sua ponta, que lhe permitem testar a consistência e estrutura dos objectos agarrados.
Em aves de rapina, o bico superior é extremamente afiado e pontiagudo, mas falta a fixação da articulação cinética, pelo que não pode produzir uma força descendente tão poderosa. Em vez disso, é usado como um instrumento de rasgar.
Em Anseriformes (a família dos patos), o bico é achatado e pode ter serrações finas nas bordas que permitem à ave filtrar partículas finas da água. Os patos como os patinhos e as pás têm este tipo de bico. Estas serrilhas podem ser desenvolvidas para uma borda recortada (por exemplo, na família das serrarias) que permite à ave agarrar alimentos escorregadios, tais como os peixes. Os anseriformes também têm terminações nervosas num prato nas pontas dos seus bicos (conhecidas como ‘pregos’) que lhes permitem encontrar alimento escondido na lama.
Em todas as aves existe uma série de ossos mais pequenos atrás dos bicos inferior e superior que lhes permitem mover o bico independentemente do crânio. Estes incluem os ossos palatino, quadrático e pterigóides e os arcos jugal. Seus movimentos exatos estão além deste texto para descrever, mas muitas das referências no final deste capítulo dão bons relatos de sua função.
Nostrils
As narinas, ou nares, estão na base do bico superior na maioria das aves e são frequentemente cercadas por uma área de pele sem penas conhecida como o cérebro. Esta pode ser muito colorida em algumas espécies, como o budgerigar, onde podem ser usadas para identificar o sexo da ave. Em muitos Anseriformes, os nares estão mais próximos da ponta do bico. As próprias nuaras são apenas aberturas para as câmaras sinusais, que por sua vez se ligam a uma rede ramificada de câmaras ósseas ao longo da cabeça da ave. Estes seios nasais variam de acordo com a espécie, mas a maioria dos pacientes aviários tem um seio infraorbital. Este fica abaixo dos olhos e está frequentemente envolvido em infecções sinusais e oculares. Difere dos seios nasais vistos na maioria dos mamíferos porque a parede lateral não tem osso, sendo coberta apenas por tecido mole. Isto significa que qualquer infecção do seio infraorbital resulta frequentemente em inchaço na face da ave ventral do olho. Estes seios nasais também comunicam com os sacos aéreos da cabeça e do pescoço. A função destes sacos aéreos não é clara, mas podem ajudar na ressonância da voz. Quando uma ave sofre de infecções nos seios nasais, as entradas estreitas destes seios podem ficar parcialmente bloqueadas e agir como válvulas unidireccionais, permitindo que o ar entre nos sacos, mas não saia. Os sacos podem então inchar excessivamente, e inchaços suaves são então comumente vistos nas costas ou na nuca da ave.
Os seios nasais e as narinas externas comunicam com a orofaringe através da fenda do cuanal. Esta é uma abertura estreita na linha média do palato duro e está localizada imediatamente sobre a glote quando o bico está fechado, permitindo que a ave respire através das narinas. É frequentemente a área escolhida para a recolha de amostras quando se tenta isolar agentes infecciosos da doença das vias aéreas superiores nas aves.
O crânio da ave liga-se ao atlas (ou primeira vértebra espinhal) através de apenas um côndilo occipital na base do crânio, ao contrário dos dois mamíferos. Há também um grande número de vértebras cervicais altamente móveis. Estes dois factores tornam a cabeça da ave extremamente ágil. Entretanto, a articulação atlanto-occipital também é um ponto fraco, tornando a luxação nesse local muito fácil.
Coluna vertebral
Vértebras cervicais
As vértebras cervicais (Figura 9.2) são independentemente móveis no paciente aviário, como são no paciente mamífero, e variam em número dependendo da espécie entre 11 e 25. São geralmente em forma de caixa.
Vértebras torácicas, lombares e sacrais
As vértebras torácicas (Figura 9.2) são fundidas em aves de rapina, pombos e muitas outras espécies para formar um único osso conhecido como o notário. Em outras espécies elas têm alguma mobilidade limitada. Existem então duas articulações intervertebrais entre o notário e as vértebras lombares e sacrais fundidas. Estas vértebras fundidas são conhecidas como o sínacro. O synsacrum funde-se com a própria pélvis para formar um escudo dorsal de osso sobre o aspecto caudal da ave.
Vértebras coccígeas
A maioria das vértebras coccígeas caudais (Figura 9.2) são geralmente fundidas em uma única estrutura conhecida como o pygostyle – que forma a parte ‘parsons nose’ da galinha!
Pelvis
O teto da pélvis é formado pelo sísacro (Figura 9.2). Os dois “lados” da pélvis são reduzidos em tamanho em relação aos mamíferos, mas consistem nos ossos ilíacos e isquiáticos, sendo o acetábulo criado onde se encontram. O acetábulo nas aves não é uma cavidade óssea completa como nos mamíferos, mas uma folha fibrosa. Existe uma crista na pelve lateral conhecida como antitrocanter, que se articula com o trocanter maior do fémur. A função desta crista é evitar que o membro seja abduzido quando empoleirado. Os ossos púbicos da pélvis não se fundem na linha média ventral como nos mamíferos. Em vez disso, eles formam ossos longos finos que se estendem caudalmente em direção ao orifício de ventilação. Eles fornecem suporte para a pele que cobre o abdômen caudal e espaço suficiente para a passagem dos ovos na ave fêmea.
Ribcage
Psittaciformes têm oito pares de costelas (Figura 9.2). Cada costela tem um segmento dorsal conhecido como costela torácica, e um segmento ventral, ou costela esternal. Estas costelas apontam para trás e ligam rigidamente as vértebras torácicas dorsalmente e a quilha, ou esterno, ventralmente.
Sternum
As vértebras esternais são fundidas em aves para formar a quilha. A quilha tem uma crista de linha média que divide os músculos peitorais em lados direito e esquerdo. A crista pode ser uma estrutura profunda, como se vê em pombos, aves de rapina e psitacídeos, permitindo a fixação de grandes músculos peitorais para um voo forte. Alternativamente, a quilha pode ser achatada, como no Anseriformes, para proporcionar uma estrutura tipo barco mais adequada à flutuação.
Asas
A articulação do ombro é formada pelo encontro de três ossos: o úmero, a escápula (que é mais tubular do que a do mamífero achatado) e um terceiro osso conhecido como coracóide (Figura 9.2). Este último osso forma uma escora que apoia a articulação do ombro contra o esterno. O músculo supracoracoide liga-se à quilha, depois passa pelo forame, ou abertura, formado no ponto de encontro destes ossos, e assim atinge o aspecto dorsal do úmero onde se liga. A contracção deste músculo, juntamente com alguns tecidos elásticos que também estão presentes, ajuda a levantar a asa. Os músculos peitorais ligam-se da quilha ao úmero para puxar a asa para baixo. As clavículas fundidas, ou osso do osso (muitas vezes chamado de furcula), articulam-se com o osso coracoide e fornecem um grau de mola para o bater das asas. O úmero é pneumonizado, o que significa que não pode ser usado para terapia com fluidos intra-ósseos. Este é também um ponto importante a considerar na reparação de fracturas.
O úmero articula-se com o rádio e ulna na articulação do cotovelo. O raio é o menor destes dois ossos e fica cranialmente. O cúbito é a fonte de fixação das penas do vôo secundário, que se inserem diretamente no periósteo deste osso (Figura 9.3). O cúbito é freqüentemente usado para administração de fluido intra-ósseo em aves.
Figure 9.3 Aspecto ventral da asa de um peneireiro (Falco tinnunculus) com as penas dissimuladas removidas mostrando a fixação dos primários ao manus e dos secundários ao cúbito.

O rádio e o ulna articulados com um osso cárpico radial e um osso cárpico ulnar, respectivamente. Estes, por sua vez, articulam-se com três metacarpos ósseos. O primeiro metacarpo ósseo é o equivalente do “polegar” aviário. É conhecido como a alula, ou “asa bastarda”, e forma uma projecção de penas a partir do aspecto craniano da articulação carpometacárpica. Os restantes dois metacarpianos são conhecidos como os metacarpianos maior e menor, e articulam-se com a primeira falange craniana e com o dígito menor caudalmente. A primeira falange articula-se então com a segunda falange, formando a ponta da asa. As penas primárias ligam-se ao periósteo das falanges e ossos metacarpianos menores (Figura 9.3).
A área da asa é aumentada por finas folhas de tecido elástico que se estendem de uma superfície articular a outra. A maior estende-se do ombro até à articulação carpal craniana e é conhecida como o propágio ou “teia da asa” (Figura 9.4). Isto pode ser usado em algumas espécies, como pombos, para administração de vacinas.
Figure 9.4 Aspecto dorsal da asa de um peneireiro (Falco tinnunculus) com as penas cobertas removidas mostrando a folha elástica do propatagium fazendo a ponte entre a articulação do cotovelo.

O membro pélvico
O acetábulo da pélvis segura a cabeça femoral (Figura 9.2). O membro pode ser bloqueado, e impedido de ser raptado, pelo trocânter maior do fémur que se envolve com a crista antitrocantérica da pélvis. O fémur é pneumonizado em muitas aves. Na articulação asfixiante, o fémur articula-se com a patela e o osso tibiotársico. O osso tibiotársico é assim chamado porque é formado a partir da fusão da tíbia e da linha proximal dos ossos tarsais, e também pode ser usado para administração de líquido intra-ósseo. No aspecto lateral da tibiotársica proximal está a fíbula muito reduzida.
Distalmente, o osso tibiotársico articula-se com o osso tarsometatarsal. Este osso é formado pela fusão da fila distal dos ossos do tarso com o osso solitário metatarso. A articulação entre o tibiotársico e o tarsometatársico é conhecida como a articulação intertarsal, ou sufrago. O tarsometatarsus articula-se então com as falanges.
Em Psittaciformes, dois dígitos apontam para a frente (o segundo e o terceiro) e dois para trás (o primeiro e o quarto), criando um membro zigodáctilo. O primeiro dígito tem duas falanges, o segundo dígito tem três falanges, o terceiro tem quatro falanges e o quarto tem cinco falanges. Nas aves empoleiradas (Passeriformes) e nas aves de rapina, o segundo, terceiro e quarto dígitos apontam para a frente e o primeiro para trás, criando um membro anisodáctilo. Algumas espécies, como a águia-pesqueira (Pandion haliaetus), podem mover o quarto dígito para frente ou para trás para ajudar a capturar sua presa, criando um membro semi-zigodáctilo.
Sentidos especiais
Olhos
O olho aviário é único, pois contém uma série de pequenos ossos. Estes são conhecidos como os ossículos esclerais (Figura 9.2). Eles formam uma estrutura em forma de anel que suporta a parte frontal do olho. O olho aviário também difere do olho de mamífero por não ser um globo, mas em forma de pêra, com a extremidade mais estreita mais externa.
O olho aviário é grande em proporção ao tamanho total do crânio, com apenas um septo ósseo fino de papel que separa as órbitas direita e esquerda. As aves têm uma terceira pálpebra móvel, translúcida, e pálpebras superiores e inferiores, sendo a inferior mais móvel do que a superior. Duas glândulas produtoras de lágrimas existem normalmente: a terceira pálpebra, ou glândula Harderian, que está localizada na base da terceira pálpebra, e a glândula lacrimal situada caudolateralmente, como nos mamíferos.
A cor do íris pode mudar com a idade em alguns papagaios, por exemplo, o papagaio cinzento africano tem um íris cinzento escuro até aos 4-5 meses de idade, quando se torna cinzento-amarelado, e depois prateado à medida que envelhece. Em outros a íris pode ser usada como indicador do sexo da ave: em grandes catatuas, por exemplo, a fêmea tem uma íris marrom-avermelhada brilhante, enquanto a do macho é escura, marrom-preto.
A retina aviária é espessa e não possui vasos sanguíneos superficiais visíveis, ao contrário da dos mamíferos. Para proporcionar nutrição à retina, as aves possuem uma estrutura vascular pregueada e dobrada chamada pecten oculi, que se encontra no ponto em que o nervo óptico entra no olho. Contrai-se intermitentemente, expelindo nutrientes para o humor vítreo.
Finalmente, a íris aviária possui fibras de músculo esquelético dentro dela, ao contrário dos mamíferos que possuem apenas fibras de músculo liso. Isto significa que o paciente aviário pode contrair e dilatar a pupila à vontade, reduzindo assim o valor do reflexo da luz pupilar como ferramenta na determinação da função ocular. Como os dois nervos ópticos estão completamente separados um do outro, o reflexo de luz consensual é também um mau indicador da função cerebral.
Ear
Não há pinna nas aves, embora algumas espécies, como as corujas de pêlo longo e curto, tenham penas nesta área. Existe um canal externo curto e horizontal, coberto por penas, que se localiza caudolateralmente à órbita ocular. A membrana timpânica pode ser vista claramente. O ouvido médio liga-se à orofaringe através do canal de Eustáquio. Os ossículos aurais dos mamíferos são substituídos na ave por uma cartilagem lateral extra de columela e um osso de columela medial que transmite ondas sonoras para o ouvido interno.
O ouvido interno contém a cóclea e os canais semicirculares, que cumprem as mesmas funções que nos mamíferos.
Anatomia respiratória
Sistema respiratório superior
As neves abrem-se para as passagens nasais, que por sua vez comunicam com a glote da laringe através de uma abertura de linha média no palato duro que forma o telhado da boca caudal. Esta abertura é chamada de fenda choanal. O sistema sinusal e os sacos aéreos cervicocefálicos foram previamente mencionados.
Laringe
Aves têm uma estrutura laríngea reduzida, faltando uma epiglote, a cartilagem da tiróide e as pregas vocais vistas em gatos e cães. A estrutura principal é a glote, que protege a entrada da traquéia. Os músculos externos puxam a glote e a traquéia para a frente, de modo que ela se comunica diretamente com a fenda do choanal, permitindo que a ave respire através de suas narinas. A glote é mantida fechada quando em repouso, abrindo-se apenas na inspiração e expiração.
Traqueia
A traqueia das espécies aviárias difere da traqueia dos mamíferos na medida em que os seus anéis de cartilagem são círculos completos, em forma de anel signet-ring, encravando um em cima do outro, em vez dos anéis em forma de C da traqueia dos mamíferos. Em Psittaciformes e aves de rapina diurnas, a forma destes anéis de cartilagem é ligeiramente achatada no sentido dorsoventral, enquanto que na maioria dos Passeriformes são redondos.
Em algumas espécies, como o cisne de Whooper e a galinha-pintada, a traquéia forma uma série de anéis e bobinas na entrada torácica. Outras espécies, como a emu, têm uma divisão ventral da linha média na traquéia a três quartos da distância entre a cabeça e a entrada do tórax. A mucosa de revestimento da traqueia projeta-se através desta fenda para formar um saco traqueal. Isto melhora a ressonância vocal. Nos patos machos, como os patinhos, existe um inchaço na última porção da traqueia, muitas vezes apenas no interior do tórax, conhecido como bula traqueal.
Syrinx
Antes de a traqueia se dividir nos dois brônquios principais, existe uma estrutura conhecida como syrinx (Figura 9.6). É aqui que a ave produz a maior parte da sua voz. Ela é composta por uma série de músculos e duas membranas que podem ser vibradas, independentemente da inspiração ou expiração.
Sistema respiratório inferior
Pulmões
Os pulmões das espécies aviárias são rígidos na estrutura e não inflamam ou esvaziam significativamente. São achatados em forma e firmemente presos ao aspecto ventral das vértebras torácicas e costelas vertebrais. Não há diafragma nas aves, e a cavidade comum do corpo é referida como o celoma.
Os brônquios pareados são suportados por anéis de cartilagem em forma de C, ao contrário da traquéia. Os brônquios primários fornecem cada um dos dois pulmões, e dividem-se rapidamente em brônquios secundários e terciários, ou parabrônquios. Existem quatro grupos principais de brônquios secundários que fornecem o pulmão, mas o seu papel na troca gasosa é mínimo. Os brônquios terciários, no entanto, desempenham um papel nas trocas gasosas, uma vez que as suas paredes estão cheias de membranas capazes de trocas gasosas. Estas áreas aparecem como pequenos fossos, ou átrios, aos quais estão ligados tubos ainda mais finos, conhecidos como capilares de ar. Estes entrelaçam-se entre si para formar uma malha tridimensional entrelaçada com os leitos capilares sanguíneos. Estes capilares de ar variam em tamanho, mas têm em média cerca de 3-5 mm de diâmetro. Este diâmetro extremamente pequeno produz forças de atracção muito elevadas entre as suas paredes quando estão presentes secreções de fluido, resultando num rápido bloqueio das superfícies respiratórias. Para evitar que isto ocorra, existem células dentro do parabrônquio que secretam o surfactante, para garantir que as vias aéreas permaneçam abertas.
A estrutura pulmonar pode ser ainda classificada pela direção do fluxo de ar dentro dela no pulmão neopulmonar e no pulmão paleopulmonar. Estes serão mencionados posteriormente quando se discutir a fisiologia respiratória.
Sacos aéreos
A parte final do sistema respiratório inferior das aves é composta pelos sacos aéreos (ver Figura 9.5). Estes são sacos semelhantes a balões que actuam como foles, bombeando o ar para dentro e para fora dos pulmões rígidos das aves em resposta aos movimentos da parede do corpo e do esterno. As paredes ar-saco são muito finas e compostas de epitélio escamoso simples que cobre uma camada de tecido conjuntivo elástico pouco vascularizado.
Figure 9.5. Sistema ar-saco aviário num pato: (a) passagens nasais; (b) seio infra-orbital; (c) sacos aéreos cervicocefálicos (simples); (d) sacos aéreos claviculares; (e) sacos aéreos torácicos cranianos; (f) sacos aéreos torácicos caudais; e (g) sacos aéreos abdominais. H, coração; L, fígado; Lu, pulmões; B, bula de seringa (patos machos).
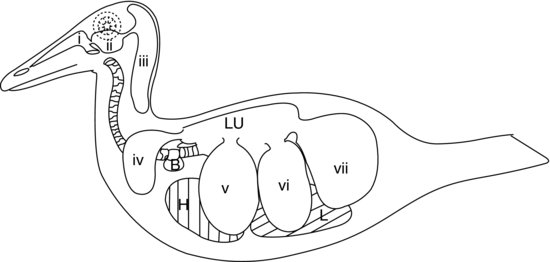
Na maioria das aves existem nove sacos aéreos. Um deles é o saco aéreo separado já mencionado, o saco aéreo cervicocefálico, que não comunica de todo com os pulmões. Os outros oito comunicam todos com os pulmões através de um brônquio secundário (excepto os sacos aéreos abdominais que se ligam ao brônquio primário de cada lado). A Figura 9.5 mostra o sistema de sacos aéreos de um pato.
Além do saco aéreo cervicocefálico separado, os outros oito sacos aéreos padrão são