Veterian Key
- De familie Anatidae, subfamilie Anatinae, bijvoorbeeld de wilde eend, de slobeend, de eidereend en de bergeend
- De familie Anatidae, subfamilie Anserinae, stam Anserini (zwanen en ganzen), bijvoorbeeld de knobbelzwaan, de wipzwaan en de bergeend, de brandgans en de grauwe gans
- De familie Falconidae, bijvoorbeeld de slechtvalk, de saker, de lanner, de giervalk
- De familie Accipitridae zoals de buizerds (gewone, ruigpoot- en honingbuizerds), de sperwer, de havik, de steenarend
De Psittaciformes behoren tot de meest kleurrijke vogels die als huisdier worden gehouden.
Nervosysteem
De hersenen van vogels zijn uiterst glad en missen de vele gyri (de ribbels in de hersenen) die bij zoogdieren voorkomen (figuur 9.1). Het gezichtsvermogen lijkt bij de meeste vogels het overheersende zintuig te zijn. Twee grote optische kwabben liggen tussen de hersenhelften en de kleine hersenen, en het is hier dat de oogzenuwen communiceren en informatie verspreiden. Er is geen callosum corpus, de hersenschors is over het algemeen zeer dun, maar het corpus striatum is goed ontwikkeld en wordt verondersteld de plaats te zijn van de mentale associatie bij vogels. Een belangrijk kenmerk van de vogelhersenen is het pijnbogenlichaam dat in het diencephalon zit, craniaal ten opzichte van de kleine hersenen in de middellijn dorsaal. Het pijnappelklichaam heeft secretiecellen die lijken op fotoreceptoren en zal dus reageren op licht. Zij zijn ook via de craniale cervicale ganglia verbonden met de oogzenuw. Het pijnappelklierorgaan is verantwoordelijk voor de regulering van vele seizoensgebonden effecten, zoals voortplanting en migratie, en voor het circadiane ritme. Het heeft een rechtstreeks effect via hormoonafscheiding op de hypothalamus.
Figuur 9.1 Dorsaal aspect van de hersenhelften van vogels toont het ontbreken van gyri.

Het zenuwstelsel van vogels is niet verschillend van dat van zoogdieren. Vogels bezitten 12 hersenzenuwen (CN), hetzelfde aantal als bij katten en honden. Bij vogels is de oogzenuw (CN II) de grootste hersenzenuw, die bijna de helft van de doorsnede van de wervelkolom heeft. Hij passeert het calvarium via één enkele opening in plaats van via meerdere kleinere die verbonden zijn met de cribriformplaat van zoogdieren. De andere 11 hersenzenuwen hebben soortgelijke functies als bij zoogdieren, en de lezer wordt verwezen naar meer diepgaande anatomische teksten voor meer informatie (King & McLelland, 1975; Bennett, 1994).
Elke vleugel heeft een zenuwvoorziening vanuit een plexus brachialis die is afgeleid van de ruggenmergzenuwen in het caudale cervicale gebied. Er zijn drie zenuwplexussen in het lumbosacrale gebied: de lumbale, de ischiatische en de pudendale. De lumbale plexus is afkomstig van de laatste twee lumbale en de eerste één tot twee sacrale ruggenmergzenuwwortels. Evenals de andere lumbosacrale plexussen ligt hij in een holte van het bekken, dorsaal van de craniale nierstreek. Hij voorziet de lichaamswand en de spieren van de bovenbenen en geeft aanleiding tot de obturatorzenuwen, de femorzenuwen, de craniale gluteale zenuwen en de venuszenuwen. In tegenstelling tot honden en katten hebben vogels een plexus ischiaticus die afkomstig is van vier tot zeven spinale zenuwen in het heiligbeengebied en die gelegen is in een holte van het bekken dorsaal van de middennierstructuur. Hier ontspringt de voornaamste zenuwvoorziening van de achterste ledematen – de nervus ischiaticus, de grootste perifere zenuw van het lichaam, en de nervus caudalis gluteus. Tenslotte vormt zich in een holte van het bekken dorsaal van de caudale nierstreek een pudendal plexus uit vijf coccygeale spinale zenuwen, die de staart en de cloacale zone innerveert.
Musculoskeletaal systeem
De meeste vogels hebben vliegvermogen. De dichte, logge botten van het aan de aarde gebonden zoogdier zouden te veel inspanning vergen om in de lucht te worden getild. Vogels hebben daarom hun skeletstructuur aangepast door het aantal botten te vereenvoudigen door sommige samen te smelten en in het algemeen de hele structuur lichter te maken door in veel botten luchtruimten te creëren.
Om het skelet nog lichter te maken, zijn verschillende van de grotere botten, en zelfs sommige wervels in de wervelkolom, direct of indirect verbonden met de luchtwegen, en men zegt dat ze gepneumoniseerd zijn. Dit vervangt de dikke medullaire holte of het beenmerg dat in het midden van de beenderen van zoogdieren aanwezig is, en levert een lichte, trabeculaire structuur op. Hoewel licht, is de structuur toch uiterst sterk.
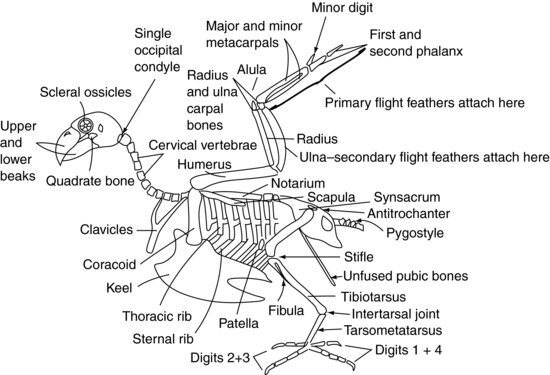
Figuur 9.2 toont een algemeen vogelskelet.
Figuur 9.2 Aviaanskelet.
Figuur 9.2 Aviaanskelet.2 Aviaanskelet.
Schedel
Snavel
De snavel, of snavel, is het belangrijkste kenmerk van de schedel van de vogels. Hij heeft een verbijsterend aantal vormen en afmetingen gekregen, voornamelijk afhankelijk van het dieet waaraan de vogel zich heeft aangepast. In alle gevallen bestaat hij uit een bovensnavel (boven- en ondersnavel) en een ondersnavel (onderkaak), die bedekt zijn met een laag keratine, een taaie eiwitverbinding die lijkt op die waaruit het exoskelet van insecten is opgebouwd. Deze keratinelaag staat bekend als de rhamphotheca. Zij wordt verder geclassificeerd zodat de bovenkaaklaag de rhinotheca wordt genoemd en de onderkaaklaag de gnatotheca. De rhinotheca en gnatotheca groeien uit een plaat aan de basis van de respectieve snavelzijden, waarbij de vervangingssnelheid afhangt van het soort voedsel dat wordt gegeten en de slijtage die de snavel ondergaat.
Bij de Psittaciformes (tabel 9.1) is de bovensnavel krachtig ontwikkeld en eindigt in een scherpe punt die over de bredere, dikkere ondersnavel uitsteekt. De enorme kracht in de snavel van een papegaai is te danken aan een synoviaal gewricht of scharniermechanisme, bekend als het kinetisch gewricht, dat de snavel met de schedel verbindt. De ondersnavel van de papegaai heeft aan de punt een reeks druksensoren, waarmee de consistentie en de structuur van de vastgegrepen voorwerpen kunnen worden getest.
Bij roofvogels is de bovensnavel uiterst scherp en puntig, maar hij heeft geen kinetisch gewricht, zodat hij niet zo’n krachtige neerwaartse kracht kan produceren. In plaats daarvan wordt hij gebruikt als een scheurinstrument.
Bij de Anseriformes (eendenfamilie) is de snavel afgeplat en kan hij aan de randen zijn voorzien van fijne kartels, waarmee de vogel fijne deeltjes uit het water kan filteren. Eenden zoals wilde eenden en slobeenden hebben dit type snavel. Deze kartels kunnen verder ontwikkeld zijn tot een gekartelde rand (b.v. bij de familie van de toepasselijk genaamde zaagbekken), waardoor de vogel zich kan vastgrijpen aan glibberig voedsel, zoals vis. Anseriformes hebben ook zenuwuiteinden in een plaat aan de punten van hun snavel (bekend als de “nagel”) waarmee ze in modder verborgen voedsel kunnen vinden.
Bij alle vogels is er een reeks kleinere beenderen achter de onder- en bovensnavel die hen in staat stellen de snavel onafhankelijk van de schedel te bewegen. Hiertoe behoren de palatijn-, quadraat- en pterygoidebeenderen en de jugaalbogen. Hun precieze bewegingen vallen buiten het bestek van deze tekst, maar veel van de referenties aan het eind van dit hoofdstuk geven een goede beschrijving van hun functie.
Nostrillen
De neusgaten, of neusgaten, liggen bij de meeste vogels aan de basis van de bovensnavel en zijn vaak omgeven door een gebied van veerloze huid dat de graan wordt genoemd. Deze kan sterk gekleurd zijn bij sommige soorten, zoals de grasparkiet, waar ze kunnen worden gebruikt om het geslacht van de vogel te identificeren. Bij veel Anseriformes liggen de neusgaten meer naar de punt van de snavel toe. De neusgaten zelf zijn slechts openingen naar de sinuskamers, die op hun beurt in verbinding staan met een vertakt netwerk van benige kamers in de gehele kop van de vogel. Deze sinussen variëren naar gelang van de soort, maar de meeste vogelpatiënten hebben een infraorbitale sinus. Deze zit onder de ogen en is vaak betrokken bij sinus- en ooginfecties. Hij verschilt van de sinussen bij de meeste zoogdieren doordat de laterale wand geen bot heeft, maar uitsluitend bedekt is met zacht weefsel. Dit betekent dat een infraorbitale sinusinfectie vaak resulteert in een zwelling op het gezicht van de vogel ventraal van het oog. Deze sinussen staan ook in verbinding met de luchtzakken in de kop en de nek. De functie van deze luchtzakken is niet duidelijk, maar ze kunnen helpen bij de resonantie van de stem. Wanneer een vogel een sinusinfectie heeft, kunnen de nauwe inlaten van deze sinussen gedeeltelijk geblokkeerd raken en als eenrichtingskleppen gaan werken, waardoor er wel lucht in maar niet uit de sinussen komt. De sinuszakken kunnen dan overmatig opblazen en zachte zwellingen zijn dan vaak te zien op de rug of de nek van de kop van de vogel.
De sinussen en de uitwendige neusgaten staan in verbinding met de orofarynx via de choanale spleet. Dit is een smalle opening in de middellijn van het harde gehemelte en bevindt zich onmiddellijk boven de glottis wanneer de snavel gesloten is, zodat de vogel door zijn neusgaten kan ademen. Het is vaak de plaats die wordt gekozen voor het nemen van monsters wanneer men besmettelijke agentia voor ziekten van de bovenste luchtwegen bij vogels probeert te isoleren.
De schedel van de vogelpatiënt staat in verbinding met de atlas (of eerste ruggenwervel) via slechts één achterhoofdsknobbel aan de basis van de schedel, in tegenstelling tot de twee bij zoogdieren. Er is ook een groot aantal zeer beweeglijke halswervels. Deze twee factoren maken de kop van de vogel uiterst beweeglijk. Het atlanto-occipitale gewricht is echter ook een zwak punt, waardoor ontwrichting op die plaats zeer gemakkelijk is.
Vertebrale kolom
Cervicale wervels
De cervicale wervels (figuur 9.2) zijn bij de aviaire patiënt onafhankelijk beweeglijk, net als bij de zoogdierpatiënt, en variëren in aantal afhankelijk van de soort tussen 11 en 25. Zij zijn over het algemeen doosvormig.
Thoracale, lumbale en sacrale wervels
De thoracale wervels (figuur 9.2) zijn bij roofvogels, duiven en vele andere soorten vergroeid tot één enkel bot, het notarium genaamd. Bij andere soorten hebben ze een beperkte beweeglijkheid. Er zijn dan twee tussenwervelgewrichten tussen het notarium en de vergroeide lenden- en heiligbeenwervels. Deze vergroeide wervels worden het synsacrum genoemd. Het synsacrum versmelt met het bekken zelf en vormt een dorsaal schild van bot over het caudale aspect van de vogel.
Coccygeale wervels
De meerderheid van de caudale coccygeale wervels (Figuur 9.2) zijn gewoonlijk vergroeid tot één enkele structuur die bekend staat als de pygostijl – die het “neuspees”-gedeelte van de kip vormt!
Pelvis
Het dak van het bekken wordt gevormd door het synsacrum (figuur 9.2). De twee “zijden” van het bekken zijn kleiner dan bij zoogdieren, maar bestaan uit het bekkenbeen en het zitbeen, waarbij het acetabulum ontstaat waar zij samenkomen. Het acetabulum is bij vogels geen volledige benige kom zoals bij zoogdieren, maar een vezelig vel. Op de zijkant van het bekken bevindt zich een kam die antitrochanter wordt genoemd en die scharniert met de trochanter van het dijbeen. De functie van deze kam is te voorkomen dat het lidmaat bij het neerkomen wordt geabduceerd. De schaambeenderen van het bekken smelten niet samen in de ventrale middellijn zoals bij zoogdieren. In plaats daarvan vormen zij fijne lange beenderen die zich caudaal uitstrekken naar de buikholte. Zij bieden steun aan de huid die het caudale achterlijf bedekt en voldoende ruimte voor de passage van eieren bij de vrouwelijke vogel.
Ribkooi
Pittaciformes hebben acht paar ribben (figuur 9.2). Elke rib heeft een dorsaal segment, de thoracale rib genoemd, en een ventraal segment, de sternale rib. Deze ribben wijzen naar achteren en vormen een stijve verbinding tussen de thoracale wervels dorsaal en de kiel, of het borstbeen, ventraal.
Sternum
De sternale wervels zijn bij vogels met elkaar vergroeid tot de kiel. De kiel heeft een middellijnrichel die de borstspieren in rechter- en linkerkant verdeelt. De kam kan diep zijn, zoals bij duiven, roofvogels en papegaaiachtigen, zodat grote borstspieren kunnen aanhechten voor een sterke vlucht. De kiel kan ook afgeplat zijn, zoals bij Anseriformes, om een bootachtige structuur te verkrijgen die meer geschikt is om te drijven.
Vleugels
Het schoudergewricht wordt gevormd door de samenkomst van drie beenderen: het opperarmbeen, het schouderblad (dat meer buisvormig is dan het afgeplatte zoogdierbot) en een derde bot dat bekend staat als het coracoid (figuur 9.2). Dit laatste bot vormt een stut die het schoudergewricht tegen het borstbeen stut. De musculus supracoracoideus hecht zich aan de kiel, gaat vervolgens door het foramen, of de opening, die op het ontmoetingspunt van deze beenderen is gevormd, en bereikt zo de dorsale zijde van het opperarmbeen, waar hij zich vasthecht. Het samentrekken van deze spier, samen met enkele elastische weefsels die ook aanwezig zijn, helpt bij het optillen van de vleugel. De borstspieren hechten zich vanuit de kiel aan het opperarmbeen vast om de vleugel naar beneden te trekken. De met elkaar vergroeide sleutelbeenderen, of vorkbeenderen (vaak furcula genoemd), articuleren met het coracoideum en zorgen voor een zekere veerkracht bij het flapperen van de vleugels. De humerus is gepneumoniseerd, wat betekent dat hij niet kan worden gebruikt voor intra-seneuze vloeistoftherapie. Dit is ook een belangrijk punt om rekening mee te houden bij het herstel van fracturen.
Het opperarmbeen articuleert met het spaakbeen en de ellepijp in het ellebooggewricht. Het spaakbeen is het kleinste van deze twee botten en ligt craniaal. De ellepijp vormt de bevestigingsbron voor de secundaire vliegveren, die rechtstreeks in het periost van dit bot grijpen (figuur 9.3). De ellepijp wordt bij vogels vaak gebruikt voor intra-osseuze vochttoediening.
Figuur 9.3 Ventraal aspect van de vleugel van een torenvalk (Falco tinnunculus) waarvan de dekveren zijn verwijderd en waarop de aanhechting van de primaire veren aan de manus en van de secundaire veren aan de ellepijp is te zien.

Het spaakbeen en de ellepijp articuleren respectievelijk met één radiaal handwortelbeen en één ulnair handwortelbeen. Deze articuleren op hun beurt met drie middenhandsbeentjes. Het eerste middenhandsbeentje is het equivalent van de aviaire ‘duim’. Het staat bekend als de alula, of “bastaardvleugel”, en vormt een vederachtige uitsteeksel van het craniale aspect van het carpometacarpale gewricht. De twee overige middenhandsbeentjes worden het grote en het kleine middenhandsbeentje genoemd, en articuleren met het eerste vingerkootje craniaal en het kleine vingerkootje caudaal. Het eerste vingerkootje scharniert dan met het tweede vingerkootje, en vormt zo de vleugeltip. De primaire veren hechten aan het periost van de vingerkootjes en de kleine middenhandsbeentjes (figuur 9.3).
Het vleugeloppervlak wordt vergroot door dunne vellen elastisch weefsel die zich van het ene gewrichtsoppervlak naar het andere uitstrekken. De grootste strekt zich uit van de schouder tot het carpale gewricht craniaal en staat bekend als het propatagium of “vleugelweb” (figuur 9.4). Dit kan bij sommige soorten, zoals duiven, worden gebruikt voor het toedienen van vaccins.
Figuur 9.4 Dorsaal aspect van de vleugel van een torenvalk (Falco tinnunculus) waarvan de dekveren zijn verwijderd, waarbij het elastische vel van het propatagium te zien is dat het ellebooggewricht overbrugt.

Bekken ledemaat
Het acetabulum van het bekken houdt de femurkop vast (figuur 9.2). De grote trochanter van het femur grijpt in de antitrochanterische kam van het bekken, waardoor het ledemaat kan worden vergrendeld en abductie wordt voorkomen. Bij veel vogels is het dijbeen gepneumoniseerd. In het gewricht van de stomp articuleert het dijbeen met de knieschijf en het tibiotarsale bot. Het tibiotarsale bot wordt zo genoemd omdat het is gevormd uit de fusie van het scheenbeen en de proximale rij tarsale botten, en kan ook worden gebruikt voor intraossale vochttoediening. Aan de laterale zijde van de proximale tibiotarsus bevindt zich de sterk gereduceerde fibula.
Distaal scharniert het tibiotarsale bot met het tarsometatarsale bot. Dit bot wordt gevormd door de fusie van de distale rij tarsale botten met het solitaire metatarsale bot. Het gewricht tussen de tibiotarsus en de tarsometatarsus wordt het intertarsale, of suffrago, gewricht genoemd. De tarsometatarsus articuleert vervolgens met de vingerkootjes.
Bij de Psittaciformes wijzen twee vingers naar voren (de tweede en de derde) en twee naar achteren (de eerste en de vierde), waardoor een zygodactyl ledemaat ontstaat. Het eerste lid heeft twee vingerkootjes, het tweede lid heeft drie vingerkootjes, het derde heeft vier vingerkootjes en het vierde heeft vijf vingerkootjes. Bij de baarsachtigen (Passeriformes) en de roofvogels wijzen de tweede, derde en vierde vinger naar voren en de eerste naar achteren, waardoor een anisodactyl ledemaat ontstaat. Bij sommige soorten, zoals de visarend (Pandion haliaetus), kan de vierde poot naar voren of naar achteren wijzen om de prooi te kunnen vangen, waardoor een semi-zygodactyl ledemaat ontstaat.
Speciale zintuigen
Oog
Het vogeloog is uniek doordat het een reeks kleine botjes bevat. Deze staan bekend als de sclerale gehoorbeentjes (figuur 9.2). Zij vormen een ringvormige structuur die de voorkant van het oog ondersteunt. Het vogeloog verschilt ook van het zoogdieroog doordat het geen oogbol is, maar peervormig, met het smallere uiteinde aan de buitenkant.
Het vogeloog is groot in verhouding tot de totale omvang van de schedel, met slechts een flinterdun benig tussenschot dat de rechter en linker oogkas van elkaar scheidt. Vogels hebben een beweeglijk, doorschijnend derde ooglid, en boven- en onderoogleden, waarvan het onderste beweeglijker is dan het bovenste. Er zijn gewoonlijk twee traanklieren: de derde oogklier, of Harderiaanse klier, die zich aan de basis van het derde ooglid bevindt, en de lacrimale klier die caudolateraal gelegen is, zoals bij zoogdieren.
De kleur van de iris kan bij sommige papegaaien met de leeftijd veranderen; zo heeft de Afrikaanse grijze papegaai een donkergrijze iris tot hij 4-5 maanden oud is, dan wordt hij geelgrijs, en vervolgens zilverkleurig naarmate hij ouder wordt. Bij andere kan de iris worden gebruikt als een indicator voor het geslacht van de vogel: bij grote kaketoes bijvoorbeeld heeft het vrouwtje een heldere, roodbruine iris, terwijl die van het mannetje donker bruinzwart is.
Het netvlies van vogels is dik en heeft geen zichtbare bloedvaten aan de oppervlakte, in tegenstelling tot dat van zoogdieren. Om het netvlies van voeding te voorzien, beschikken vogels over een geplooide en geplooide vaatstructuur, de pecten oculi, die zich bevindt op het punt waar de oogzenuw het oog binnenkomt. Deze trekt met tussenpozen samen, waardoor voedingsstoffen in het glasvocht terechtkomen.
Ten slotte heeft de iris van vogels skeletspiervezels in zich, in tegenstelling tot zoogdieren die alleen gladde spiervezels hebben. Dit betekent dat de aviaire patiënt zijn pupil naar believen kan vernauwen en verwijden, waardoor de waarde van de pupilreflex als instrument voor het bepalen van de oogfunctie afneemt. Omdat de twee oogzenuwen volledig van elkaar gescheiden zijn, is de consensuele lichtreflex ook een slechte indicator van de hersenfunctie.
oor
Er is geen pinna bij vogels, hoewel sommige soorten, zoals de ransuil en de velduil, veren hebben in dit gebied. Er is een kort, horizontaal uitwendig kanaal, bedekt met veren, dat zich caudolateraal van de oogkas bevindt. Het trommelvlies kan duidelijk worden gezien. Het middenoor staat in verbinding met de oropharynx via het kanaal van Eustachius. De gehoorbeentjes van zoogdieren zijn bij de vogel vervangen door een lateraal, extra columella-kraakbeen en een mediaal columella-botje, die geluidsgolven doorgeven aan het binnenoor.
Het binnenoor bevat het slakkenhuis en de halfcirkelvormige kanalen, die dezelfde functies vervullen als bij zoogdieren.
Anatomie van de ademhaling
Bovenste ademhalingsstelsel
De neusgaten komen uit in de neusholten, die op hun beurt in verbinding staan met de glottis van het strottenhoofd via een opening in de middellijn in het harde gehemelte dat het dak van de caudale mond vormt. Deze opening wordt de choanale spleet genoemd. Het sinusstelsel en de cervicocephale luchtzakken zijn reeds eerder vermeld.
Larynx
Vogels hebben een gereduceerde larynxstructuur, zonder epiglottis, schildkraakbeen en stemplooien, zoals bij katten en honden. De belangrijkste structuur is de glottis, die de toegang tot de luchtpijp beschermt. Externe spieren trekken het strotklepje en de luchtpijp naar voren, zodat deze in directe verbinding staat met de keelholte, waardoor de vogel door zijn neusgaten kan ademen. De glottis wordt in rust gesloten gehouden en gaat alleen open bij inademing en uitademing.
Trachea
De trachea van vogelsoorten verschilt van de trachea van zoogdieren doordat de kraakbeenringen volledige, zegelringvormige cirkels zijn, die boven elkaar in elkaar grijpen, in plaats van de C-vormige ringen van de trachea van zoogdieren. Bij de Psittaciformes en de dagactieve roofvogels is de vorm van deze kraakbeenringen lichtjes afgeplat in dorsoventrale richting, terwijl zij bij de meeste Passeriformes rond zijn.
Bij sommige soorten, zoals de Whooper’s zwaan en de parelhoen, vormt de luchtpijp een reeks lussen en spiralen aan de thoracale inham. Bij andere soorten, zoals de emoe, is de luchtpijp op driekwart van de afstand tussen de kop en de borstholte ventraal gespleten. Het slijmvlies van de tracheale bekleding steekt door deze spleet en vormt zo een tracheale zak. Dit verbetert de resonantie van de stem. Bij mannelijke eenden, zoals wilde eenden, is er een zwelling in het laatste deel van de luchtpijp, vaak net binnen de borstkas, die bekend staat als de tracheale bulla.
Syrinx
Voordat de luchtpijp zich splitst in de twee hoofdbronchiën, is er een structuur die bekend staat als de syrinx (figuur 9.6). Dit is de plaats waar de vogel het grootste deel van zijn stem produceert. Hij bestaat uit een reeks spieren en twee membranen die, onafhankelijk van inspiratie of uitademing, in trilling kunnen worden gebracht.
Lager ademhalingssysteem
Longen
De longen van vogelsoorten zijn star van structuur en blazen niet sterk op of leeg. Ze zijn afgeplat van vorm en stevig bevestigd aan het ventrale aspect van de borstwervels en de wervelribben. Er is geen middenrif bij vogels, en de gemeenschappelijke lichaamsholte wordt de coelom genoemd.
De gepaarde bronchiën worden ondersteund door C-vormige ringen van kraakbeen, in tegenstelling tot de trachea. De primaire bronchiën bevoorraden elk van de twee longen, en splitsen zich snel in secundaire en tertiaire bronchiën, of parabronchi. Er zijn vier hoofdgroepen van secundaire bronchiën die de longen bevoorraden, maar hun rol in de gasuitwisseling is minimaal. De tertiaire bronchiën spelen echter wel een rol bij de gasuitwisseling, omdat hun wanden gevuld zijn met membranen die in staat zijn tot gasuitwisseling. Deze gebieden zien eruit als kleine putjes, of atria, waaraan nog fijnere buisjes zijn verbonden die bekend staan als luchtcapillairen. Deze verstrengelen zich met elkaar tot een driedimensionaal netwerk dat verweven is met de haarvatenbedden in het bloed. Deze luchtcapillairen variëren in grootte, maar hebben gemiddeld een diameter van 3-5 mm. Deze uiterst kleine diameter veroorzaakt bij vloeibare afscheidingen zeer grote aantrekkingskrachten tussen hun wanden, hetgeen leidt tot een snelle verstopping van de ademhalingsoppervlakken. Om dit te voorkomen zijn er cellen in de parabronchi die surfactant afscheiden, zodat de luchtwegen open blijven.
De longstructuur kan verder worden ingedeeld naar de richting van de luchtstroming in de long in de neopulmonische long en de paleopulmonische long. Deze zullen later worden genoemd bij de bespreking van de ademhalingsfysiologie.
Luchtzakken
Het laatste deel van het onderste ademhalingsstelsel van vogels bestaat uit de luchtzakken (zie figuur 9.5). Dit zijn ballonachtige zakjes die als blaasbalgen fungeren en de lucht in en uit de stijve vogellongen pompen als reactie op bewegingen van de lichaamswand en het borstbeen. De wanden van de luchtzakjes zijn zeer dun en bestaan uit eenvoudig plaveiselepitheel dat een laag slecht gevasculariseerd elastisch bindweefsel bedekt.
Figuur 9.5 Aviaire lucht-sac-systeem in een eend: (a) neusgangen; (b) infra-orbitale sinus; (c) cervicocephale luchtzakken (enkel); (d) claviculaire luchtzakken; (e) craniale thoracale luchtzakken; (f) caudale thoracale luchtzakken; en (g) abdominale luchtzakken. H, hart; L, lever; Lu, longen; B, syringeale bulla (mannelijke eenden).
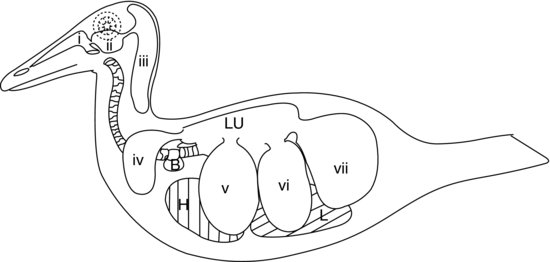
Bij de meeste vogels zijn er negen luchtzakken. Eén daarvan is de reeds genoemde afzonderlijke luchtzak, de cervicocephale luchtzak, die in het geheel niet met de longen in verbinding staat. De andere acht communiceren alle met de longen via een secundaire bronchus (behalve de abdominale luchtzakken die aan elke kant met de primaire bronchus in verbinding staan). Figuur 9.5 toont het luchtzaksysteem van een eend.
Naast de afzonderlijke cervicocefale luchtzak zijn de andere standaard acht luchtzakken