Chiave veterinaria
- La famiglia Anatidae, sottofamiglia Anatinae, per esempio l’anatra domestica, il germano reale, l’edredone e la volpoca
- La famiglia Anatidae, sottofamiglia Anserinae, Tribù Anserini (cigni e oche vere), per esempio i cigni muti, Whooper e Bewick, le oche barbagianni e greylag
- La famiglia Falconidae per esempio il falco pellegrino, il saker, il lanario, il girfalco
- La famiglia Accipitridae come le poiane (comune, dalle zampe ruvide e miele), lo sparviero, l’astore, l’aquila reale
I Psittaciformes sono tra i più colorati uccelli tenuti come animali domestici.
Sistema nervoso
Il cervello aviario è estremamente liscio, privo dei molti giroscopi (le creste del cervello) visti nei mammiferi (Figura 9.1). La vista sembra essere il senso dominante nella maggior parte degli uccelli. Due grandi lobi ottici si trovano tra gli emisferi cerebrali e il cervelletto, ed è qui che i nervi ottici comunicano e diffondono le informazioni. Non c’è corpo calloso, la corteccia cerebrale è generalmente molto sottile, ma il corpo striato è ben sviluppato e si pensa che sia il sito di associazione mentale negli uccelli. Una caratteristica importante del cervello degli uccelli è il corpo pineale che si trova nel diencefalo, craniale al cervelletto in linea mediana dorsale. Il corpo pineale ha cellule secretorie simili ai fotorecettori e quindi risponde alla luce. Sono collegati anche attraverso i gangli cervicali cranici al nervo ottico. Il corpo pineale è responsabile della regolazione di molti effetti stagionali come la riproduzione e la migrazione e i ritmi circadiani. Ha un effetto diretto attraverso la secrezione di ormoni sull’ipotalamo.
Figura 9.1 Aspetto dorsale degli emisferi cerebrali aviari che mostra la mancanza di gyri.

Il sistema nervoso aviario non è dissimile da quello visto nella sua controparte mammifera. Gli uccelli possiedono 12 nervi cranici (CN), lo stesso numero che nei gatti e nei cani. Negli uccelli, il nervo ottico (CN II) è il più grande nervo cranico, essendo quasi la metà del diametro della colonna vertebrale. Passa attraverso il calvario attraverso un unico foro piuttosto che attraverso più piccoli associati alla placca cribrosa dei mammiferi. Gli altri 11 nervi cranici hanno funzioni simili a quelle dei mammiferi, e si rimanda il lettore a testi di anatomia più approfonditi per maggiori informazioni (King & McLelland, 1975; Bennett, 1994).
Ognuna delle ali ha un apporto nervoso da un plesso brachiale derivato dai nervi spinali nella zona cervicale caudale. Ci sono tre plessi nervosi nella regione lombosacrale: lombare, ischiatico e pudendo. Il plesso lombare deriva dalle ultime due radici nervose lombari e dalle prime due sacrali. Come gli altri plessi lombosacrali, si trova in una cavità del bacino, dorsalmente alla zona del rene craniale. Rifornisce la parete del corpo e i muscoli della parte superiore delle gambe e dà origine ai nervi otturatore, femorale, gluteo craniale e safena. A differenza di cani e gatti, gli uccelli hanno un plesso ischiatico che deriva da quattro a sette nervi spinali nella zona sacrale e che si trova in una cavità del bacino dorsale alla struttura del rene medio. Dà origine al principale apporto nervoso per gli arti posteriori – il nervo ischiatico che è il più grande nervo periferico del corpo e il nervo gluteo caudale. Infine, un plesso pudendo si forma in una cavità del bacino dorsale alla zona caudale del rene da cinque nervi spinali coccigei e innerva la coda e la zona cloacale.
Sistema muscoloscheletrico
La maggior parte degli uccelli ha il potere di volare. Le ossa dense e ingombranti dei mammiferi legati alla terra richiederebbero uno sforzo eccessivo per sollevarsi in aria. Gli uccelli hanno quindi adattato la loro struttura scheletrica, semplificando il numero di ossa fondendone alcune insieme, e generalmente alleggerendo l’intera struttura creando spazi d’aria all’interno di molte delle ossa.
Per alleggerire ulteriormente lo scheletro, molte delle ossa più grandi, e anche alcune delle vertebre della colonna vertebrale, sono collegate direttamente o indirettamente alle vie aeree, e sono dette pneumonizzate. Questo sostituisce la spessa cavità midollare o midollo osseo presente al centro delle ossa dei mammiferi, e produce una struttura leggera e trabecolare. Pur essendo leggera, la struttura è tuttavia estremamente forte.
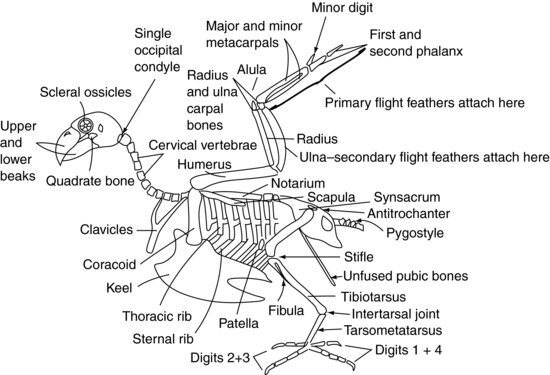
La figura 9.2 mostra uno scheletro aviario generalizzato.
Figura 9.2 Scheletro aviario.
Cranio
Beak
Il becco, o bill, è la caratteristica principale del cranio aviario. È stato modificato in un numero sconcertante di forme e dimensioni, a seconda della dieta a cui l’uccello si è adattato. In tutti i casi è composto da un becco superiore (mascellare) e inferiore (mandibolare) che sono coperti da uno strato di cheratina, un composto proteico duro simile a quello che forma l’esoscheletro degli insetti. Questo strato di cheratina è noto come rhamphotheca. È ulteriormente classificato in modo che lo strato mascellare è indicato come rinoteca, e lo strato mandibolare come gnatoteca. La rinoteca e la gnatoteca crescono da una placca alla base dei rispettivi lati del becco, il tasso di sostituzione dipende dal tipo di cibo mangiato e dall’abrasione che il becco riceve.
Negli Psittaciformes (Tabella 9.1), il becco superiore è potentemente sviluppato e termina con una punta affilata che sovrasta il più largo e robusto becco inferiore. L’enorme potenza del becco di un pappagallo è dovuta ad un’articolazione sinoviale o meccanismo a cerniera, noto come articolazione cinetica, che unisce il becco al cranio. Il becco inferiore del pappagallo ha una serie di sensori di pressione sulla sua punta, che gli permettono di testare la consistenza e la struttura degli oggetti afferrati.
Nei rapaci, il becco superiore è estremamente affilato e appuntito, ma manca l’attacco del giunto cinetico, quindi non può produrre una forza verso il basso così potente. Invece, è usato come uno strumento di strappo.
Negli Anseriformes (la famiglia delle anatre), il becco è appiattito e può avere sottili dentellature ai bordi che permettono all’uccello di filtrare particelle fini dall’acqua. Le anatre come i germani e le spatole hanno questo tipo di becco. Queste dentellature possono essere ulteriormente sviluppate in un bordo frastagliato (per esempio nella famiglia chiamata giustamente sawbill) che permette all’uccello di afferrare il cibo scivoloso, come il pesce. Gli anseriformi hanno anche terminazioni nervose in una piastra sulla punta del becco (conosciuta come “unghia”) che permette loro di trovare il cibo nascosto nel fango.
In tutti gli uccelli c’è una serie di piccole ossa dietro il becco inferiore e superiore che permettono loro di muovere il becco indipendentemente dal cranio. Questi includono le ossa palatine, quadrate e pterigoidi e le arcate jugal. I loro movimenti esatti non possono essere descritti in questo testo, ma molti dei riferimenti alla fine di questo capitolo danno buoni resoconti della loro funzione.
Nostre
Le narici, o narici, si trovano alla base del becco superiore nella maggior parte degli uccelli e sono spesso circondate da una zona di pelle senza piume nota come cervello. Questo può essere molto colorato in alcune specie, come il pappagallino, dove può essere usato per identificare il sesso dell’uccello. In molti Anseriformi le narici si trovano più verso la punta del becco. Le narici stesse sono semplicemente delle aperture nelle camere dei seni, che a loro volta si collegano con una rete ramificata di camere ossee in tutta la testa dell’uccello. Questi seni variano a seconda della specie, ma la maggior parte dei pazienti aviari ha un seno infraorbitale. Questo si trova sotto gli occhi ed è spesso coinvolto in infezioni sinusali e oculari. Si differenzia dai seni visti nella maggior parte dei mammiferi in quanto la parete laterale non ha osso, essendo coperta solo da tessuto molle. Questo significa che qualsiasi infezione del seno infraorbitale si traduce spesso in un gonfiore sulla faccia dell’uccello ventrale all’occhio. Questi seni comunicano anche con i sacchi d’aria della testa e del collo. La funzione di queste sacche d’aria non è chiara, ma possono aiutare la risonanza della voce. Quando un uccello soffre di infezioni sinusali, gli stretti ingressi di questi seni possono essere parzialmente bloccati e agire come valvole a senso unico, permettendo all’aria di entrare nei sacchi ma non di uscire. I sacchi possono quindi gonfiarsi eccessivamente, e gonfiori morbidi si vedono comunemente sulla schiena o sulla nuca dell’uccello.
I seni e le narici esterne comunicano con l’orofaringe attraverso la fessura choanale. Questa è una stretta apertura nella linea mediana del palato duro e si trova immediatamente sopra la glottide quando il becco è chiuso, permettendo all’uccello di respirare attraverso le narici. È spesso l’area scelta per prelevare campioni quando si cerca di isolare agenti infettivi per le malattie delle vie aeree superiori negli uccelli.
Il cranio del paziente aviario si collega all’atlante (o prima vertebra spinale) attraverso un solo condilo occipitale alla base del cranio, a differenza dei due dei mammiferi. Ci sono anche un gran numero di vertebre cervicali molto mobili. Questi due fattori rendono la testa aviaria estremamente agile. Tuttavia, l’articolazione atlanto-occipitale è anche un punto debole, rendendo molto facile la dislocazione in quel sito.
Colonna vertebrale
Vertebre cervicali
Le vertebre cervicali (Figura 9.2) sono indipendentemente mobili nel paziente aviario, come lo sono nel paziente mammifero, e variano in numero a seconda della specie tra 11 e 25. Sono generalmente di forma scatolare.
Vertebre toraciche, lombari e sacrali
Le vertebre toraciche (Figura 9.2) sono fuse nei rapaci, nei piccioni e in molte altre specie per formare un unico osso noto come notarium. In altre specie hanno una mobilità limitata. Ci sono poi due articolazioni intervertebrali tra il notarium e le vertebre lombari e sacrali fuse. Queste vertebre fuse sono conosciute come synsacrum. Il sinsacro si fonde con il bacino stesso per formare uno scudo dorsale di osso sopra l’aspetto caudale dell’uccello.
Vertebre coccigee
La maggior parte delle vertebre coccigee caudali (Figura 9.2) sono di solito fuse in un’unica struttura conosciuta come il pigostilo – che forma la parte “naso di parsons” del pollo!
Pelvis
Il tetto del bacino è formato dal sinsacro (Figura 9.2). I due “lati” del bacino sono di dimensioni ridotte rispetto ai mammiferi, ma sono costituiti dalle ossa iliache e ischiatiche, e l’acetabolo si crea dove si incontrano. L’acetabolo negli uccelli non è una presa ossea completa come nei mammiferi, ma un foglio fibroso. C’è una cresta sul bacino laterale conosciuta come antitrocantere, che si articola con il grande trocantere del femore. La funzione di questa cresta è di evitare che l’arto sia abdotto quando si appollaia. Le ossa pubiche del bacino non si fondono nella linea mediana ventrale come nei mammiferi. Formano invece delle sottili ossa lunghe che si estendono caudalmente verso lo sfiatatoio. Forniscono supporto alla pelle che copre l’addome caudale e spazio sufficiente per il passaggio delle uova nell’uccello femmina.
Ribbia
Gli psittaciformi hanno otto paia di costole (Figura 9.2). Ogni costola ha un segmento dorsale conosciuto come costola toracica, e un segmento ventrale, o costola sternale. Queste costole puntano all’indietro e collegano rigidamente le vertebre toraciche dorsalmente e la chiglia, o sterno, ventralmente.
Sterno
Le vertebre sternali sono fuse negli uccelli per formare la chiglia. La chiglia ha una linea mediana che divide i muscoli pettorali in destra e sinistra. La cresta può essere una struttura profonda, come si vede nei piccioni, nei rapaci e nei Psittaciformes, permettendo ai grandi muscoli pettorali di attaccarsi per un volo forte. In alternativa, la chiglia può essere appiattita, come negli Anseriformi, per fornire una struttura simile ad una barca più adatta al galleggiamento.
Alle
L’articolazione della spalla è formata dall’incontro di tre ossa: l’omero, la scapola (che è più tubolare di quella appiattita dei mammiferi) e un terzo osso noto come il coracoide (Figura 9.2). Quest’ultimo osso forma un puntello che sostiene l’articolazione della spalla contro lo sterno. Il muscolo sopracoracoideo si attacca alla chiglia, poi passa attraverso il forame, o apertura, formato nel punto di incontro di queste ossa, e così raggiunge la faccia dorsale dell’omero dove si attacca. La contrazione di questo muscolo, insieme ad alcuni tessuti elastici anch’essi presenti, aiuta a sollevare l’ala. I muscoli pettorali si attaccano dalla chiglia all’omero per tirare l’ala verso il basso. Le clavicole fuse, o wishbone (spesso chiamato furcula), si articolano con l’osso coracoide e forniscono un certo grado di molla al battito delle ali. L’omero è pneumonizzato, il che significa che non può essere usato per la terapia con fluidi intraossei. Questo è anche un punto importante da considerare quando si riparano le fratture.
L’omero si articola con il radio e l’ulna all’articolazione del gomito. Il radio è la più piccola di queste due ossa e si trova cranialmente. L’ulna fornisce la fonte di attacco per le penne secondarie di volo, che si inseriscono direttamente nel periostio di questo osso (Figura 9.3). L’ulna è spesso usata per la somministrazione di fluidi intraossei negli uccelli.
Figura 9.3 Aspetto ventrale dell’ala di un gheppio (Falco tinnunculus) con le piume di copertura rimosse che mostra l’attacco delle primarie alla manus e le secondarie all’ulna.

Il radio e l’ulna si articolano rispettivamente con un osso carpale radiale e uno ulnare. Questi a loro volta si articolano con tre ossa metacarpali. Il primo osso metacarpale è l’equivalente del “pollice” aviario. È conosciuto come l’alula, o “ala bastarda”, e forma una proiezione piumata dall’aspetto craniale dell’articolazione carpometacarpale. Le altre due ossa metacarpali sono conosciute come le ossa metacarpali maggiore e minore, e si articolano con la prima falange cranialmente e con il dito minore caudalmente. La prima falange si articola poi con la seconda falange, formando la punta dell’ala. Le penne primarie si attaccano al periostio delle falangi e delle ossa metacarpali minori (Figura 9.3).
L’area dell’ala è ampliata da sottili fogli di tessuto elastico che vanno da una superficie articolare all’altra. Il più grande si estende dalla spalla all’articolazione carpale cranialmente ed è conosciuto come il propatagio o “rete alare” (Figura 9.4). Questo può essere usato in alcune specie, come i piccioni, per la somministrazione di vaccini.
Figura 9.4 Aspetto dorsale dell’ala di un gheppio (Falco tinnunculus) con le piume di copertura rimosse, mostrando il foglio elastico del propatagio che collega l’articolazione del gomito.

Arto pelvico
L’acetabolo del bacino tiene la testa del femore (Figura 9.2). L’arto può essere bloccato, e impedito dall’abduzione, dal maggiore trocantere del femore che si impegna con la cresta antitrocanterica sul bacino. Il femore è pneumonizzato in molti uccelli. All’articolazione della zampa, il femore si articola con la rotula e l’osso tibiotarsico. L’osso tibiotarsico è così chiamato perché è formato dalla fusione della tibia e della fila prossimale di ossa tarsali, e può anche essere usato per la somministrazione di fluidi intraossei. Sull’aspetto laterale del tibiotarsus prossimale si trova il perone molto ridotto.
Distintamente, l’osso tibiotarsico si articola con l’osso tarsometatarsale. Questo osso è formato dalla fusione della fila distale delle ossa tarsali con l’osso metatarsale solitario. L’articolazione tra il tibiotarsus e il tarsometatarsus è nota come articolazione intertarsale o suffrago. Il tarsometatarso si articola poi con le falangi.
Nei Psittaciformes, due dita puntano in avanti (la seconda e la terza) e due indietro (la prima e la quarta), creando un arto zigodattilo. Il primo dito ha due falangi, il secondo dito ha tre falangi, il terzo ha quattro falangi e il quarto ha cinque falangi. Negli uccelli posatoi (Passeriformi) e nei rapaci, la seconda, la terza e la quarta cifra puntano in avanti e la prima punta all’indietro creando un arto anisodattilo. Alcune specie, come il falco pescatore (Pandion haliaetus), possono spostare il quarto dito in avanti o indietro per aiutare la cattura della preda, creando un arto semi-zigodattilo.
Sensi speciali
Occhio
L’occhio aviario è unico in quanto contiene una serie di piccole ossa. Questi sono conosciuti come ossicini sclerali (Figura 9.2). Essi formano una struttura ad anello che sostiene la parte anteriore dell’occhio. L’occhio aviario differisce anche dall’occhio dei mammiferi in quanto non è un globo, ma a forma di pera, con l’estremità più stretta all’esterno.
L’occhio aviario è grande in proporzione alla dimensione complessiva del cranio, con solo un setto osseo sottile come la carta che separa le orbite destra e sinistra. Gli uccelli hanno una terza palpebra mobile e traslucida, e palpebre superiori e inferiori, di cui la inferiore è più mobile della superiore. Esistono comunemente due ghiandole che producono lacrime: la terza palpebra, o ghiandola Harderiana, che si trova alla base della terza palpebra, e la ghiandola lacrimale situata caudolateralmente, come nei mammiferi.
Il colore dell’iride può cambiare con l’età in alcuni pappagalli, per esempio il pappagallo grigio africano ha un’iride grigio scuro fino a 4-5 mesi di età, quando diventa giallo-grigio, e poi argento con il passare degli anni. In altri l’iride può essere usata come indicatore del sesso dell’uccello: nei grandi cacatua, per esempio, la femmina ha un’iride brillante, rosso-marrone, mentre quella del maschio è scura, marrone-nera.
La retina aviaria è spessa e non possiede vasi sanguigni superficiali visibili, diversamente da quella dei mammiferi. Per fornire nutrimento alla retina, gli uccelli possiedono una struttura vascolare pieghettata e piegata chiamata pecten oculi, che si trova nel punto in cui il nervo ottico entra nell’occhio. Si contrae a intermittenza, espellendo le sostanze nutritive nell’umore vitreo.
Infine, l’iride aviaria ha fibre muscolari scheletriche al suo interno, a differenza dei mammiferi che hanno solo fibre muscolari lisce. Questo significa che il paziente aviario può restringere e dilatare la pupilla a suo piacimento, riducendo così il valore del riflesso della luce pupillare come strumento per determinare la funzione oculare. Poiché i due nervi ottici sono completamente separati l’uno dall’altro, il riflesso luminoso consensuale è anche un povero indicatore della funzione cerebrale.
Orecchio
Non c’è pinna negli uccelli, anche se alcune specie, come i gufi dalle orecchie lunghe e corte, hanno piume in questa zona. C’è un breve canale esterno orizzontale, coperto da piume, che si trova caudolaterale all’orbita oculare. La membrana timpanica può essere vista chiaramente. L’orecchio medio si collega all’orofaringe attraverso il canale di Eustachio. Gli ossicini uditivi dei mammiferi sono sostituiti nell’uccello da una cartilagine laterale extra columella e da un osso columella mediale che trasmettono le onde sonore all’orecchio interno.
L’orecchio interno contiene la coclea e i canali semicircolari, che svolgono le stesse funzioni dei mammiferi.
Anatomia respiratoria
Sistema respiratorio superiore
Le narici si aprono nelle fosse nasali, che a loro volta comunicano con la glottide della laringe attraverso un’apertura mediana nel palato duro che forma il tetto della bocca caudale. Questa apertura è chiamata fessura choanale. Il sistema dei seni e i sacchi d’aria cervicocefalici sono stati precedentemente menzionati.
Laringe
Gli uccelli hanno una struttura laringea ridotta, mancando l’epiglottide, la cartilagine tiroidea e le pliche vocali che si vedono nei gatti e nei cani. La struttura principale è la glottide, che protegge l’ingresso della trachea. I muscoli esterni tirano la glottide e la trachea in avanti in modo che comunichi direttamente con la fessura choanale, permettendo all’uccello di respirare attraverso le narici. La glottide è tenuta chiusa a riposo, aprendosi solo durante l’inspirazione e l’espirazione.
Trachea
La trachea delle specie aviarie differisce da quella dei mammiferi in quanto i suoi anelli cartilaginei sono cerchi completi, a forma di anello con sigillo, incastrati uno sull’altro, piuttosto che gli anelli a forma di C della trachea dei mammiferi. Nei Psittaciformes e nei rapaci diurni, la forma di questi anelli cartilaginei è leggermente appiattita in direzione dorsoventrale, mentre nella maggior parte dei Passeriformi sono rotondi.
In alcune specie, come il cigno selvatico e la faraona, la trachea forma una serie di anelli e spire all’ingresso toracico. Altre specie, come l’emu, hanno una scissione ventrale sulla linea mediana della trachea a tre quarti della distanza tra la testa e l’ingresso toracico. La mucosa di rivestimento tracheale si proietta attraverso questa fessura per formare un sacco tracheale. Questo migliora la risonanza vocale. Nelle anatre maschio, come i germani reali, c’è un rigonfiamento nell’ultima porzione della trachea, spesso proprio all’interno del torace, noto come bolla tracheale.
Siringa
Prima che la trachea si divida nei due bronchi principali, c’è una struttura conosciuta come la siringa (Figura 9.6). È qui che l’uccello produce la maggior parte della sua voce. È composta da una serie di muscoli e due membrane che possono essere fatte vibrare, indipendentemente dall’inspirazione o dall’espirazione.
Sistema respiratorio inferiore
Polmoni
I polmoni delle specie aviarie sono rigidi nella struttura e non si gonfiano o sgonfiano significativamente. Sono di forma appiattita e saldamente attaccati all’aspetto ventrale delle vertebre toraciche e delle costole vertebrali. Non c’è diaframma negli uccelli, e la cavità corporea comune è indicata come il celoma.
I bronchi appaiati sono sostenuti da anelli di cartilagine a forma di C, a differenza della trachea. I bronchi primari riforniscono ciascuno dei due polmoni e si dividono rapidamente in bronchi secondari e terziari, o parabronchi. Ci sono quattro gruppi principali di bronchi secondari che riforniscono il polmone, ma il loro ruolo nello scambio di gas è minimo. I bronchi terziari, tuttavia, giocano un ruolo nello scambio di gas, poiché le loro pareti sono piene di membrane capaci di scambio gassoso. Queste aree appaiono come piccole fosse, o atri, a cui sono collegati tubi ancora più sottili noti come capillari d’aria. Questi si intrecciano tra loro per formare una rete tridimensionale intrecciata con i letti capillari del sangue. Questi capillari d’aria variano in dimensione, ma in media circa 3-5 mm di diametro. Questo diametro estremamente piccolo produce forze di attrazione molto elevate tra le loro pareti quando sono presenti secrezioni fluide, con conseguente rapido blocco delle superfici respiratorie. Per evitare che questo accada, ci sono cellule all’interno dei parabronchi che secernono surfattante, per garantire che le vie aeree rimangano aperte.
La struttura del polmone può essere ulteriormente classificata in base alla direzione del flusso d’aria al suo interno nel polmone neopolmonare e nel polmone paleopolmonare. Questi saranno menzionati più avanti quando si parlerà di fisiologia respiratoria.
I sacchi d’aria
La parte finale del sistema respiratorio inferiore aviario è composta dai sacchi d’aria (vedi Figura 9.5). Questi sono sacchi simili a palloncini che agiscono come mantici, pompando l’aria dentro e fuori i rigidi polmoni aviari in risposta ai movimenti della parete del corpo e dello sterno. Le pareti dei sacchi aerei sono molto sottili e composte da un semplice epitelio squamoso che copre uno strato di tessuto connettivo elastico scarsamente vascolarizzato.
Figura 9.5 Sistema aviario dei sacchi aerei in un’anatra: (a) passaggi nasali; (b) seno infra-orbitale; (c) sacchi aerei cervicocefalici (singoli); (d) sacchi aerei clavicolari; (e) sacchi aerei toracici craniali; (f) sacchi aerei toracici caudali; e (g) sacchi aerei addominali. H, cuore; L, fegato; Lu, polmoni; B, bolla siringea (anatre maschio).
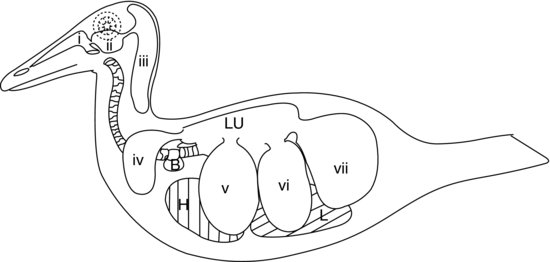
Nella maggioranza degli uccelli ci sono nove sacchi d’aria. Uno di questi è il sacco d’aria separato già menzionato, il sacco d’aria cervicocefalico, che non comunica affatto con i polmoni. Gli altri otto comunicano tutti con i polmoni attraverso un bronco secondario (tranne i sacchi d’aria addominali che si collegano al bronco primario su ogni lato). La figura 9.5 mostra il sistema dei sacchi d’aria di un’anatra.
Oltre al sacco d’aria cervicocefalico separato, gli altri otto sacchi d’aria standard sono