Frontiers in Cellularand Infection Microbiology
Introduction
Bacteroides fragilis, qui se trouve dans la flore gastro-intestinale des humains et du bétail, est une bactérie anaérobie. C’est l’un des commensaux humains proéminents et l’un des Bacteroides isolés les plus courants des échantillons cliniques qui causent la diarrhée, la péritonite, les abcès intra-abdominaux, la septicémie et les infections purulentes endogènes (Sears et al…, 1995, 2014).
Il a été démontré que B. fragilis prévient les maladies inflammatoires intestinales chez l’animal atteint de colite en raison de la production d’une molécule immunomodulatrice, le polysaccharide A (PSA), qui induit une réponse immunitaire anti-inflammatoire dans le tissu intestinal (Mazmanian et al., 2008 ; Lee et al., 2018). Le pouvoir pathogène de B. fragilis est dû à plusieurs facteurs, notamment la capsule, les protéines de la membrane externe (OMP) et des enzymes spéciales qui comprennent une métalloprotéase de 20 kDa appelée toxine de Bacteroides fragilis (BFT) (Sears et al, 1995, 2014).
La B. fragilis productrice de BFT, la B. fragilis entérotoxinogène (ETBF), est connue comme une cause de troubles diarrhéiques chez les humains et les animaux (Myers et al., 1987 ; Purcell et al., 2017). ETBF est connu comme un facteur de risque pour les maladies inflammatoires de l’intestin (MICI) et il est présent dans les selles et les échantillons de biopsie des patients (Prindiville et al., 2000 ; Basset et al., 2004 ; Zamani et al., 2017). L’expression de BFT est révélée pour cliver le domaine extracellulaire de la E-cadhérine, qui est un constituant structurel majeur de la zonule d’adhérence et est responsable de l’adhésion cellulaire. De plus, le BFT peut activer la signalisation de la B-caténine et induire la sécrétion d’IL8 dans les cellules épithéliales du côlon (Wu et al., 1998). Une étude a indiqué que suite au traitement des cellules HT29/C1 par le BFT, la perte de l’E-cadhérine associée à la membrane a initié la localisation nucléaire de la ß-caténine, ce qui a induit la traduction de c-myc et a conduit à une prolifération cellulaire persistante (Wu et al., 2003). La puissance de l’ETBF et son influence sur la structure et la physiologie de l’épithélium gastro-intestinal suggèrent que la présence de l’ETBF pourrait contribuer aux maladies coliques chroniques, y compris la transformation oncogène, l’inflammation intestinale, les dysfonctionnements coliques chroniques et les lésions précancéreuses et cancéreuses colorectales (Wu et al, 1998, 2003 ; Sears et al., 2014).
Le cancer colorectal (CCR) est l’un des cancers les plus répandus dans le monde et représente 9 % de tous les cancers et il est la quatrième cause de décès liés au cancer dans le monde (Sears et al., 1995 ; Ferlay et al., 2015). Cependant, il existe une différence significative entre les pays en ce qui concerne l’incidence standardisée par âge de ce cancer ; le taux d’incidence aux États-Unis et dans les pays européens est plus de 25 fois supérieur à celui des pays africains et asiatiques. Le taux de morbidité et de mortalité du CCR a diminué ces dernières années en raison de l’amélioration des tests de dépistage qui peuvent détecter les modifications colorectales à des stades plus précoces et de l’amélioration des médicaments et des procédures (Boyle et Langman, 2000 ; Rafiemanesh et al…, 2016).
Le CCR est l’un des cancers les plus fréquents en Iran et constitue le troisième cancer le plus fréquent chez les hommes iraniens (8,1-8,3 pour 100 000 populations) et le quatrième cancer le plus répandu chez les femmes iraniennes (6,5-7,5 pour 100 000 populations) (Moghimi-Dehkordi et al., 2008 ; Kolahdoozan et al., 2010). Les facteurs de risque du CCR sont l’obésité, la sédentarité, un régime riche en graisses, un régime pauvre en légumes et en fruits, le tabagisme, l’abus d’alcool et la prise d’anti-inflammatoires non stéroïdiens (AINS). L’inflammation chronique, les MICI, les polypes, les adénomes et la dysplasie provoquent des modifications des cellules du côlon et les rendent sujettes au cancer (Boyle et Langman, 2000 ; Johnson et al., 2013).
L’incidence et la mortalité du CCR diminuent dans les nations occidentales développées, tandis que son incidence augmente chez les deux sexes au cours des dernières décennies en Iran en raison des changements de mode de vie et de régime alimentaire. Une autre raison de cette diminution peut être l’augmentation du nombre d’installations et l’amélioration de l’équipement et de la technologie, car plus de personnes se réfèrent aux établissements de soins de santé pour le dépistage, alors que dans le passé, une personne aurait pu avoir un cancer et même être décédée, mais le cancer n’a pas été diagnostiqué en raison du manque d’équipement et d’installations (Malekzadeh et al., 2009 ; Siegel et al…, 2014).
Dans cette étude, la fréquence et l’abondance de l’ETBF dans les échantillons de biopsie des patients atteints de CCR et de conditions précancéreuses ont été comparées à celles des individus sans antécédents personnels ou familiaux de maladie colorectale afin d’étudier l’association entre la présence de l’ETBF et le développement tumoral.
Matériels et méthodes
Patients et spécimens
Dans cette étude cas-témoins, 120 biopsies de la muqueuse ont été collectées chez des patients iraniens avec une condition précancéreuse et CRC (n = 68) et dans le groupe de contrôle (n = 52) en utilisant la coloscopie. Les patients présentant des lésions précancéreuses (lésions dentelées, adénomes, dysplasie de bas grade : LGD et dysplasie de haut grade : HGD) et cancéreux (cancer colorectal : CRC) qui ont été référés à l’hôpital Imam Khomeini de Téhéran entre mars 2015 et janvier 2017 ont été sélectionnés pour participer à cette étude. Le comité d’éthique de l’université des sciences médicales de Téhéran a approuvé le protocole de l’étude. De plus, le consentement éclairé a été obtenu de tous les participants. Tous les patients ont été diagnostiqués sur la base des symptômes cliniques ainsi que des normes histologiques et radiographiques, qui ont montré des caractéristiques typiques avec une distribution particulière (Swiderska et al., 2014). Toutes les données sur l’âge, le sexe et le type de lésions ont été extraites des dossiers des patients. Tous les patients qui entrent dans l’étude ont été diagnostiqués au moment de la coloscopie et la chimiothérapie n’a pas commencé à les traiter.
Pendant la même période, 52 témoins sains, sans antécédents personnels ou familiaux de maladie colorectale diagnostiquée, dont l’âge et le sexe correspondaient à ceux des patients ont été inclus dans l’étude comme témoins. Des antécédents récents de diarrhée et de MICI constituaient un critère d’exclusion pour les témoins. Aucune des personnes ayant participé à cette étude n’a utilisé d’antibiotiques ou de probiotiques au cours des 3 derniers mois. Tous les spécimens ont été conservés dans un récipient stérile contenant du milieu thioglycollate (Merck, Allemagne) et transportés au laboratoire dans des conditions anaérobies pour être immédiatement manipulés. En outre, 2 biopsies muqueuses ont été prélevées sur chaque patient pour la culture et l’extraction d’ADN.
Culture bactérienne
Deux homogénéisateurs en verre ont été utilisés pour la perturbation mécanique et l’homogénéisation de toutes les biopsies ; ensuite, elles ont été cultivées sur un milieu Bacteroides Bile Esculin agar (BBE) (Himedia Laboratories Pvt. Ltd, Inde). Dans cette étude, le système Anoxomat (MART Microbiology Drachten, Pays-Bas) a été utilisé pour fournir des conditions atmosphériques gazeuses pour les anaérobies ; ensuite, les plaques ont été incubées dans une chambre anaérobie à 37°C pendant 72 h. De plus, B. fragilis a été confirmé par la méthode PCR en temps réel.
Extraction d’ADN
L’ADN a été extrait directement du tissu de la biopsie en utilisant le kit RTP® Mycobacteria (Invitek, Berlin, Allemagne). La densité optique (OD) de l’ADN extrait a été déterminée à 260 nanomètres. Ensuite, l’ADN a été conservé à -20°C pour les analyses ultérieures et la PCR en temps réel.
PCR en temps réel
Les séquences du gène bft et de l’ARNr 16S ont été récupérées de la banque de gènes. Les amorces et les sondes ont été conçues en utilisant l’amorce 3 plus (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). Toutes les amorces et sondes utilisées pour la détection de tous les types de gènes bft avaient été conçues dans notre étude précédente (Zamani et al., 2017). Afin de détecter les sous-types de gènes bft, la PCR en temps réel a été réalisée comme décrit précédemment (Merino et al., 2011).
L’ADN génomique de la souche ETBF D-134 et du clone 1 de RIGLD ont été utilisés comme témoins positifs (Rashidan et al., 2018). Le contrôle négatif était le mélange maître PCR TaqMan avec de l’eau distillée à la place de l’ADN.
L’ADN standard a été préparé pour l’amplification et le nombre de molécules de la matrice par gramme a été calculé par la formule définie (Zamani et al., 2017). La courbe standard pour le bft et le gène de l’ARNr 16S a été évaluée en utilisant chaque amorce et sonde avec une dilution sérielle de 10 fois des échantillons d’ADN de B. fragilis, correspondant à 101-106 valeur moyenne par gramme de biopsies.
Selon la courbe standard et l’ordonnée à l’origine, les échantillons qui n’ont pas montré le signal fluorescent avant le Ct de 38 ont été déterminés comme négatifs. De même, les échantillons qui ont produit une fluorescence d’une valeur Ct ≤ 10 ont été dilués. L’efficacité de la PCR en temps réel a été déterminée comme E = 10(-1/slop) – 1 (Zamani et al., 2017).
Après optimisation des courbes standard, la série de dilution a été mise dans chaque run d’amplification. Les tests de PCR en temps réel ont été évalués à l’aide de l’outil de PCR en temps réel LinGene K (Bioer, Hangzhou, PR, Chine). Tous les tests ont été réalisés dans un volume de 25 μL (Zamani et al., 2017). Pour garantir la qualité, toutes les expériences ont été répétées une deuxième fois indépendamment et les moyennes ont été rapportées.
Pour vérifier la spécificité de la PCR et la taille attendue du produit, les amorces ont été appliquées dans une PCR classique et les amplicons ont été passés sur le gel d’agarose. De plus, la spécificité des fragments amplifiés positifs a été prouvée par séquençage. Le produit de la PCR a été envoyé à la Macrogen Corporation de Corée du Sud pour le séquençage. Les résultats du séquençage ont été analysés avec le logiciel Chromas 2.6. Ensuite, toutes les séquences ont été analysées dans la base de données NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Dans tous les isolats séquencés, le pourcentage de similitude avec les gènes apparentés (gène de l’ARNr 16s et gène bft) présents dans la base de données NCBI était supérieur à 95%.
Analyse statistique
Les résultats ont été comparés à l’aide des tests exacts de Fisher, du Chi carré et de Mann-Whitney. P < 0,05 a été considéré comme statistiquement significatif. De même, les valeurs moyennes ± l’erreur-type de la moyenne (SEM) ont été calculées pour B. fragilis et l’ETBF. Les données ont été analysées à l’aide du logiciel statistique SPSS 13.0.
Dans cette étude, la régression logistique conditionnelle a été utilisée pour évaluer les odds ratios (OR) et les intervalles de confiance à 95%. Dans ce modèle, l’odds ratio est un estimateur cohérent du ratio de taux de CCR lors de l’exposition à l’ETBF par rapport aux sujets non exposés.
Résultats
Dans cette étude, 68 échantillons ont été prélevés chez des patients atteints de CCR (36 hommes et 32 femmes ; âge moyen : 55 ans ; plage : 35-78) et 52 échantillons ont été prélevés chez des témoins sains (HC) (30 hommes et 22 femmes ; âge moyen : 56 ans ; plage : 42-78). Tous les patients ont été diagnostiqués sur la base de procédures de coloscopie au moment de l’échantillonnage. Tous les patients ont été diagnostiqués au moment de la coloscopie, ce qui donne un total de 26 patients avec un CCR invasif, 18 patients avec des lésions dentelées, 24 patients avec un adénome, 14 patients avec un LGD et 10 patients avec un HGD selon la classification actuelle (Figure 1). Ces biopsies ont été recueillies du côté droit (ascendant), du côté gauche (descendant) et des deux côtés chez 22, 32 et 14 patients, respectivement (tableau 1).
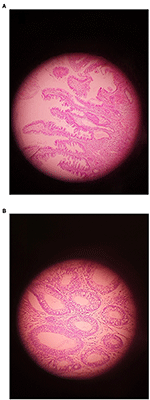
Figure 1. Un exemple d’adénome montrant (A) LGD et (B) HGD (grossissement original ×400).
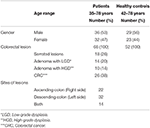
Tableau 1. Caractéristiques des patients et des HC.
Les résultats de la culture de B. fragilis étaient positifs pour 31(51%) et 25 (48%) échantillons des patients et des contrôles sains, respectivement (tableau 2) (P = 0,7).

Tableau 2. Données sur la culture de Bacteroides fragilis.
Selon la courbe standard, les dilutions de l’ADN de l’ETBF (contrôle positif) à 101, 102, 103, 104, 105 et 106 ont fourni des valeurs Ct de 15,08 ± 0,1, 18,04 ± 0,2, 21,42 ± 0,2, 25,02 ± 0,2, 28,02 ± 0,4 et 31,96 ± 0,4, respectivement. L’efficacité de la PCR en temps réel était comprise entre 98 et 100%.
Les échantillons positifs pour le gène de l’ARNr 16S et le gène bft étaient de 63 et 47% chez les patients. Cependant, des résultats positifs ont été montrés dans 81 et 3,8% des échantillons HC, respectivement (Tableau 3). La différence entre la positivité du gène bft chez les patients et les HC était statistiquement significative (P = 0,00).
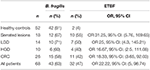
Tableau 3. Le nombre d’échantillons positifs pour le gène de l’ARNr 16S et les gènes bft dans les groupes clinicopathologiques et HC.
De même, le nombre d’échantillons positifs pour les gènes bft dans le CRC et HC au sein des groupes clinicopathologiques est présenté dans le tableau 3. L’OR le plus élevé a été trouvé dans le groupe des lésions dentelées, suivi des lésions adénomateuses avec le groupe LGD. De même, l’OR pour tous les patients (OR 22,22, IC 95 % : 5, 98,74) décrivait l’association de l’ETBF et de l’existence de lésions.
Les résultats de l’analyse quantitative de la PCR en temps réel pour le gène de l’ARNr 16S compté pour B. fragilis et le gène bft compté pour l’ETBF par ng d’ADN sont présentés dans le tableau 4. La différence entre les nombres de copies du gène de l’ARNr 16S chez les patients et les HC n’était pas statistiquement significative (P ≥ 0,05). Le nombre de copies du gène bft était plus important dans les échantillons des patients que dans ceux des témoins sains (P = 0,00). De plus, le nombre de copies du gène bft était plus important dans les échantillons positifs à l’ETBF dans le groupe des lésions précurseurs par rapport à ceux atteints de CCR ; cependant, cette différence n’était pas statistiquement significative. Les résultats du séquençage ont confirmé la présence des gènes bft et 16S rRNA (fiche technique 1).
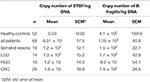
Tableau 4. Analyse quantitative du gène de l’ARNr 16S et des gènes bft dans les groupes clinicopathologiques et HC.
De plus, les résultats de ces échantillons ont indiqué que le sous-type le plus répandu du gène bft était le bft1 suivi du bft2. Sur les 32 isolats porteurs du gène bft, 18 (56,2%) isolats ont été sous-typés bft-1 et 14 (43,7%) isolats ont été sous-typés bft-2. En outre, le gène bft a été détecté dans 2 des échantillons de HC, et tous ont été sous-typés comme bft-1. Aucun échantillon d’ADN n’hébergeait le sous-type bft-3.
Discussion
Il existe des preuves croissantes de l’effet de la dysbiose microbienne dans l’intestin, de l’initiation et du développement du cancer colorectal (Sears et Garrett, 2014 ; Gagniere et al., 2016). Bien que l’incidence du CCR ait été signalée comme étant plus faible en Iran par rapport à d’autres pays, il est prévu que son taux augmente à l’avenir (Hosseini et al., 2004 ; Malekzadeh et al., 2009). Dans des études antérieures, il a également été signalé que l’ETBF peut avoir un rôle dans la diarrhée et les MICI (Myers et al., 1987 ; Prindiville et al., 2000 ; Basset et al., 2004 ; Merino et al., 2011 ; Purcell et al., 2017 ; Zamani et al, 2017).
Certaines enquêtes ont suggéré que certaines espèces bactériennes (par exemple, ETBF) peuvent fonctionner comme des bactéries pathogènes qui clarifient le développement d’une dysbiose dans la communauté microbienne de l’intestin et déclenchent le CRC (Hajishengallis et al., 2012 ; Hajishengallis et Lamont, 2016). La tumorigenèse colorectale a été induite par les réponses immunitaires et l’activation des cytokines pro-inflammatoires dues au BFT (Wu et al., 2004, 2009).
Les résultats de cette étude ont indiqué que l’ETBF était significativement associé aux lésions dentelées suivies par le LGD. Le rapport de cotes plus élevé dans ces lésions a également montré que l’exposition à l’ETBF est un facteur de risque pour les états cancéreux et surtout précancéreux. Ceci soutient l’hypothèse que les souches productrices d’ETBF peuvent jouer un rôle important dans le déclenchement de l’inflammation et de la réponse immunologique chez les personnes génétiquement sensibles et peuvent conduire au CCR. La première étude démontrant une prévalence accrue d’ETBF dans les échantillons de selles de patients atteints de cancer colorectal (38%) par rapport au groupe témoin (12%) a été menée par Ulger Toprak et al. (2006). En outre, certains rapports antérieurs ont montré une association entre le gène bft et le CCR, en particulier au stade avancé du CCR (Dejea et al., 2014 ; Boleij et al., 2015 ; Viljoen et al., 2015). Une étude menée par Purcell et al. a démontré des associations significatives de l’ETBF avec les adénomes tubulaires, les lésions dentelées et la dysplasie de bas grade, ce qui était similaire aux résultats de la présente étude (Purcell et al., 2017).
Dans cette étude, les lésions précurseurs, y compris celles avec une dysplasie de bas grade, ont montré une tendance à l’augmentation de la quantité de gène bft par rapport à celles avec un CRC ; cependant, cette différence n’était pas statistiquement significative. De plus, les auteurs ont précédemment rapporté que les marqueurs ETBF étaient observés dans le côlon et l’iléon terminal des patients atteints de colite ulcéreuse qui sont prédisposés au CCR (Zamani et al., 2017 ; Rashidan et al., 2018). De même, dans une étude précédente, seul le sous-type bft1 de ce gène a été détecté (Zamani et al., 2017). Bien que le gène bft1 soit le sous-type le plus répandu, le gène bft2 a également été trouvé dans cette étude.
Dans cette étude, l’ETBF a été détecté dans les lésions des patients atteints de CCR, et des résultats similaires ont également été rapportés précédemment par d’autres chercheurs qui ont étudié l’ETBF dans des échantillons de la muqueuse colique (Boleij et al., 2015 ; Viljoen et al., 2015). Des études récentes sur le rôle du microbiome intestinal ont démontré que la dysbiose de la communauté microbienne se produit dans les régions non tumorales et tumorales des patients atteints de CCR (Dejea et al., 2014 ; Flemer et al., 2017 ; Purcell et al., 2017). Dans ces études ont montré que certaines des bactéries, y compris Fusobacterium nucleatum, ETBF, Escherichia coli, Streptococcus gallolyticus et Enterococcus faecalis et les bactéries productrices de butyrate peuvent jouer des rôles importants dans le développement du CRC (Dejea et al, 2014 ; Flemer et al., 2017 ; Park et al., 2018).
Les résultats de la PCR en temps réel pour la détection de B. fragilis ont montré que plus de pourcentage des patients et des HC contenaient B. fragilis par rapport à la méthode de culture. La méthode de référence pour la détection des bactéries est la culture, mais elle nécessite un nombre élevé de cellules viables et, en particulier pour la culture de bactéries anaérobies, certaines limitations peuvent se produire. Peut-être que ces limitations affectent la différence entre les résultats.
Les différences dans les cultures de B. fragilis et les résultats de la PCR en temps réel entre les contrôles et les échantillons de CRC ne sont pas significatives. Ces données suggèrent que probablement les souches qui hébergent le gène bft, et non B. fragilis par lui-même, pourraient contribuer au CRC dans cette étude. Il a été démontré que B. fragilis est un commensal humain important, cette bactérie peut donc être isolée chez les témoins sains comme chez les patients. Mais chez les patients présentant des lésions cancéreuses et précancéreuses, certaines des souches qui contiennent les gènes bft ont augmenté et cette dysbiose peut probablement induire une inflammation. En outre, certaines études ont montré que le commensal humain B. fragilis a empêché le développement de la colite et peut fournir une stratégie thérapeutique efficace pour le CRC, tandis que diverses études suggèrent que les souches entérotoxigènes de cette bactérie sont associées à des tumeurs intestinales en raison de la production d’entérotoxine (Sears et al., 1995, 2014 ; Mazmanian et al., 2008 ; Lee et al., 2018).
Il y avait certaines limites dans cette étude. Différentes méthodes ont été utilisées pour diagnostiquer le gène bft, qui a été extrait directement des tissus tumoraux dans le côlon des patients. Il est donc fortement recommandé de mener une vaste étude de cohorte basée sur la population. En outre, des enquêtes supplémentaires sont nécessaires pour prouver une corrélation possible entre la présence du gène bft et les lésions dentelées, le LGD et le CRC.
Conclusions
Les résultats de cette étude suggèrent que l’ETBF pourrait être présent dans les biopsies muqueuses des patients présentant des conditions précancéreuses, telles que les lésions dentelées et le LGD, en plus des patients atteints de CRC. Une association significative a été trouvée entre la présence de l’ETBF dans les tissus affectés et le nombre de ces bactéries dans les échantillons des patients, en particulier dans les lésions précancéreuses carcinogènes : adénomes avec dysplasie de bas grade et lésions dentelées. En fait, l’ETBF est plus souvent détecté dans les lésions précoces, mais des recherches supplémentaires avec un nombre plus élevé de spécimens semblent utiles pour le déterminer précisément.
Des recherches supplémentaires sont également nécessaires pour déterminer si l’âge, le sexe, le régime alimentaire et d’autres facteurs environnementaux affectent le diagnostic de l’ETBF chez l’homme au fil du temps.
Enfin, l’ETBF pourrait être un marqueur du pronostic du CCR, en particulier dans les lésions précancéreuses, et pourrait être utilisé pour dépister ces troubles.
Data Availability Statement
Les données brutes soutenant les conclusions de ce manuscrit seront mises à disposition par les auteurs, sans réserve indue, à tout chercheur qualifié.
Ethics Statement
Les études impliquant des participants humains ont été examinées et approuvées par le Comité d’éthique de l’Université des sciences médicales de Téhéran. Les patients/participants ont fourni leur consentement éclairé écrit pour participer à cette étude.
Contributions des auteurs
SZ est le premier auteur qui a réalisé toutes les expériences de laboratoire, collecté et analysé les données, et rédigé le manuscrit. RT est le gastro-entérologue qui a effectué la coloscopie et fourni les échantillons de tous les cas. LS a participé à la conception et à la coordination de l’étude et a donné des conseils dans toutes les parties de l’étude. AS et SJ ont participé à la collecte des échantillons et à la réalisation des tests. MF a supervisé toutes les parties de l’étude. Tous les auteurs ont lu et approuvé le manuscrit.
Financement
Ce travail a été soutenu par l’Université des sciences médicales de Téhéran (subvention n° : 32467).
Conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de toute relation commerciale ou financière qui pourrait être interprétée comme un conflit d’intérêts potentiel.
Remerciements
Les auteurs tiennent à remercier le personnel de l’hôpital Imam Khomeini pour son aimable soutien dans la collecte des échantillons.
Matériel supplémentaire
Le matériel supplémentaire pour cet article peut être trouvé en ligne à : https://www.frontiersin.org/articles/10.3389/fcimb.2019.00449/full#supplementary-material
Fiche de données 1. Les résultats du séquençage des gènes bft et 16S rRNA dans un isolat.
Basset, C., Holton, J., Bazeos, A., Vaira, D., et Bloom, S. (2004). Les espèces Helicobacter et les Bacteroides fragilis entérotoxigènes sont-ils impliqués dans les maladies intestinales in?ammatoires ? Dig. Dis. Sci. 49, 1425-1432. doi : 10.1023/B:DDAS.0000042241.13489.88
CrossRef Full Text | Google Scholar
Boleij, A., Hechenbleikner, E. M., Goodwin, A. C., Badani, R., Stein, E. M., Lazarev, M. G., et al. (2015). Le gène de la toxine de Bacteroides fragilis est prévalent dans la muqueuse du côlon des patients atteints de cancer colorectal. Clin. Infect. Dis. 60, 208-215. doi : 10.1093/cid/ciu787
PubMed Abstract | CrossRef Full Text | Google Scholar
Boyle, P., et Langman, J. (2000). ABC du cancer colorectal : épidémiologie. BMJ 321, 805-808. doi : 10.1136/bmj.321.7264.805
PubMed Abstract | CrossRef Full Text | Google Scholar
Dejea, C. M., Wick, E. C., Hechenbleikner, E. M., White, J. R., Mark Welch, J. L., Rossetti, B. J., et al. (2014). L’organisation du microbiote est une caractéristique distincte des cancers colorectaux proximaux. Proc. Natl. Acad. Sci. U.S.A. 111, 18321-18326. doi : 10.1073/pnas.1406199111
PubMed Abstract | CrossRef Full Text | Google Scholar
Ferlay, J., Soerjomataram, I., Dikshit, R., Eser, S., Mathers, C., Rebelo, M., et al. (2015). Incidence du cancer et mortalité dans le monde : sources, méthodes et principaux modèles dans GLOBOCAN 2012. Int. J. Cancer 136, E359-E386. doi : 10.1002/ijc.29210
PubMed Abstract | CrossRef Full Text | Google Scholar
Flemer, B., Lynch, D. B., Brown, J. M., Jeffery, I. B., Ryan, F. J., Claesson, M. J., et al. (2017). Microbiote associé à la tumeur et non associé à la tumeur dans le cancer colorectal. Gut 66, 633-643. doi : 10.1136/gutjnl-2015-309595
PubMed Abstract | CrossRef Full Text | Google Scholar
Gagniere, J., Raisch, J., Veziant, J., Barnich, N., Bonnet, R., Buc, E., et al. (2016). Déséquilibre du microbiote intestinal et cancer colorectal. World J. Gastroenterol. 22, 501-518. doi : 10.3748/wjg.v22.i2.501
PubMed Abstract | CrossRef Full Text | Google Scholar
Hajishengallis, G., Darveau, R. P., et Curtis, M. A. (2012). The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 10, 717-725. doi : 10.1038/nrmicro2873
PubMed Abstract | CrossRef Full Text | Google Scholar
Hajishengallis, G., et Lamont, R. J. (2016). Danser avec les étoiles : comment les interactions bactériennes chorégraphiées dictent la nososymbiocité et donnent naissance à des pathogènes clés de voûte, des pathogènes accessoires et des pathobiontes. Trends Microbiol. 24, 477-489. doi : 10.1016/j.tim.2016.02.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Hosseini, S. V., Izadpanah, A., et Yarmohammadi, H. (2004). Changements épidémiologiques dans le cancer colorectal à Shiraz, Iran : 1980-2000. ANZ J. Surg. 74, 547-549. doi : 10.1111/j.1445-2197.2004.03064.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Johnson, C. M., Wei, C., Ensor, J. E., Smolenski, D. J., Amos, C. I., Levin, B., et al. (2013). Méta-analyses des facteurs de risque du cancer colorectal. Cancer Causes Control. 24:120722. doi : 10.1007/s10552-013-0201-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Kolahdoozan, S., Sadjadi, A., Radmard, A. R., et Khademi, H. (2010). Cinq cancers communs en Iran. Arch. Iran Med. 13, 143-146.
PubMed Abstract | Google Scholar
Lee, Y. K., Mehrabian, P., Boyajian, S., Wu, W. L., Selicha, J., Vonderfecht, S., et al. (2018). Le rôle protecteur de bacteroides fragilis dans un modèle murin de cancer colorectal associé à la colite. mSphere. 3:e00587-e00518. doi : 10.1128/mSphere.00587-18
PubMed Abstract | CrossRef Full Text | Google Scholar
Malekzadeh, R., Bishehsari, F., Mahdavinia, M., et Ansari, R. (2009). Épidémiologie et génétique moléculaire du cancer colorectal en Iran : une revue. Arch. Iran Med. 12:161-169.
PubMed Abstract | Google Scholar
Mazmanian, S. K., Round, J. L., et Kasper, D. L. (2008). Un facteur de symbiose microbienne prévient les maladies inflammatoires intestinales. Nature. 453, 620-625. doi : 10.1038/nature07008
PubMed Abstract | CrossRef Full Text | Google Scholar
Merino, V. R., Nakano, V., Liu, C., Song, Y., Finegold, S. M., et Avila-Campos, M. J. (2011). Détection quantitative des sous-types de Bacteroides fragilis entérotoxigènes isolés chez des enfants avec et sans diarrhée. J. Clin. Microbiol. 49, 416-418. doi : 10.1128/JCM.01556-10
PubMed Abstract | CrossRef Full Text | Google Scholar
Moghimi-Dehkordi, B., Safaee, A., et Zali, M. (2008). Facteurs pronostiques chez 1 138 patients iraniens atteints de cancer colorectal. Int. J. Colorect. Dis. 23, 683-688. doi : 10.1007/s00384-008-0463-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, L. L., Shoop, D. S., Stackhouse, L. L., Newman, F. S., Flaherty, R. J., Letson, G. W., et al. (1987). Isolation of enterotoxigenic Bacteroides fragilis from humans with diarrhoea. J. Clin. Microbiol. 25, 2230-2233.
Google Scholar
Park, C. H., Eun, C. S., et Han, D. S. (2018). Microbiote intestinal, inflammation chronique et cancer colorectal. Intest. Res. 16:338. doi : 10.5217/ir.2018.16.3.338
PubMed Abstract | CrossRef Full Text | Google Scholar
Prindiville, T., Sheikh, R., Cohen, S., Tang, Y., Cantrell, M., et Silva, J. (2000). Séquences du gène de l’entérotoxine de Bacteroides fragilis chez les patients atteints de maladie intestinale inflammatoire. Emerg. Infect. Dis. 6, 171-174. doi : 10.3201/eid0602.000210
CrossRef Full Text | Google Scholar
Purcell, R. V., Pearson, J., Aitchison, A., Dixon, L., Frizelle, F. A., et Keenan, J. I. (2017). La colonisation avec Bacteroides fragilis entérotoxigène est associée à la néoplasie colorectale à un stade précoce. PLoS ONE 12:e0171602. doi : 10.1371/journal.pone.0171602
PubMed Abstract | CrossRef Full Text | Google Scholar
Rafiemanesh, H., Pakzad, R., Abedi, M., Kor, Y., Moludi, J., Towhidi, F., et al. (2016). Cancer colorectal en Iran : tendances épidémiologiques et morphologiques. EXCLI J. 15, 738-744. doi : 10.17179/2Fexcli2016-346
PubMed Abstract | CrossRef Full Text | Google Scholar
Rashidan, M., Azimirad, M., Alebouyeh, M., Ghobakhlou, M., Aghdaei, H. A., et Zali, M. R. (2018). Détection du groupe B. fragilis et diversité de l’entérotoxine bft et des marqueurs de résistance aux antibiotiques cepA, cfiA et nim parmi les souches intestinales de Bacteroides fragilis chez les patients atteints de maladies inflammatoires de l’intestin. Anaerobe 50, 93-100. doi : 10.1016/j.anaerobe.2018.02.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Sears, C. L., et Garrett, W. S. (2014). Les microbes, le microbiote et le cancer du côlon. Cell Host Microbe 15, 317-328. doi : 10.1016/j.chom.2014.02.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Sears, C. L., Geis, A. L., et Housseau, F. (2014). Bacteroides fragilis subvertit la biologie des muqueuses : du symbiote à la carcinogenèse du côlon. J. Clin. Invest. 124, 4166-4172. doi : 10.1172/JCI72334
PubMed Abstract | CrossRef Full Text | Google Scholar
Sears, C. L., Myers, L. L., Lozenby, A., and Van Tassel, R. L. (1995). Bacteroides fragilis entérotoxigène. Clin Infect Dis. 20(suppl):142-148. doi : 10.1093/clinids/20.Supplement_2.S142
CrossRef Full Text | Google Scholar
Siegel, R., DeSantis, C., et Jemal, A. (2014). Statistiques sur le cancer colorectal, 2014. Cancer J. Clinic. 64, 104-117. doi : 10.3322/caac.21220
PubMed Abstract | CrossRef Full Text | Google Scholar
Swiderska, M., Choromanska, B., Dabrowska, E., Konarzewska-Duchnowska, E., Choromanska, K., Szczurko, G., et al. (2014). Le diagnostic du cancer colorectal. Contempor Oncol. 18, 1-6. doi : 10.5114/wo.2013.39995
PubMed Abstract | CrossRef Full Text | Google Scholar
Ulger Toprak, N., Yagci, A., Gulluoglu, B. M., Akin, M. L., Demirkalem, P., Celenk, T., et al. (2006). A possible role of Bacteroides fragilis enterotoxin in the aetiology of colorectal cancer. Clin. Microbiol. Infect. 12, 782-786. doi : 10.1111/j.1469-0691.2006.01494.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Viljoen, K. S., Dakshinamurthy, A., Goldberg, P., et Blackburn, J. M. (2015). Le profilage quantitatif des bactéries associées au cancer colorectal révèle des associations entre fusobacterium spp, Bacteroides fragilis entérotoxigène (ETBF) et les caractéristiques clinicopathologiques du cancer colorectal. PLoS ONE 10:e0119462. doi : 10.1371/journal.pone.0119462
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, S., Lim, K. C., Huang, J., Saidi, R. F., et Sears, C. L. (1998). L’entérotoxine de Bacteroides fragilis clive la protéine d’adhérence de la zonule, E-cadhérine. Proc. Natl. Acad. Sci. U.S.A. 95, 14979-14984. doi : 10.1073/pnas.95.25.14979
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, S., Morin, P. J., Mauyo, D. J. I. K., et Sears, C. (2003). L’entérotoxine de Bacteroides fragilis induit l’expression de c-myc et la prolifération cellulaire. Gastroenterology 124, 392-400. doi : 10.1053/gast.2003.50047
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, S., Powell, J., Mathioudakis, N., Kane, S., Fernandez, E., et Sears, C. L. (2004). L’entérotoxine de Bacteroides fragilis induit la sécrétion d’interleukine-8 par les cellules épithéliales intestinales par l’intermédiaire de protéines kinases activées par des agents mitogènes et d’une voie du facteur nucléaire kappaB régulée par la tyrosine kinase. Infect. Immun. 72, 5832-5839. doi : 10.1128/IAI.72.10.5832-5839.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, S., Rhee, K. J., Albesiano, E., Rabizadeh, S., Wu, X., Yen, H. R., et al. (2009). Un commensal colique humain favorise la tumorigenèse du côlon via l’activation des réponses des cellules T helper de type 17. Nat. Med. 15, 1016-1022. doi : 10.1038/nm.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Zamani, S., Shariati, S. H., Zali, M. R., Aghdaei, H. A., Asiabar, A. S., Bokaie, S., et al. (2017). Détection de Bacteroides fragilis entérotoxiques chez les patients atteints de colite ulcéreuse. Gut Pathogens 9:53. doi : 10.1186/s13099-017-0202-0
PubMed Abstract | CrossRef Full Text | Google Scholar
.