Veterian Key
- Familien Anatidae, underfamilie Anatinae, f.eks. gråand, spadeand, edderfugl og gråand
- Familien Anatidae, underfamilie Anserinae, Stamme Anserini (svane og sædgæs), f.eks. stumme svaner, sangsvaner og Bewick-svaner, bramgæs og grågæs
- Falconidae-familien f.eks. vandrefalke, saker, llanner, jagtfalken
- Familien Accipitridae som f.eks. musvågen (musvåge, råbuk og honningfalk), spurvehøg, spurvehøg, kongeørn
Psittaciformes er blandt de mest farvestrålende fugle, der holdes som kæledyr.
Nervesystem
Fuglenes hjerne er ekstremt glat og mangler de mange gyri (de riller i hjernen), som ses hos pattedyr (figur 9.1). Synet synes at være den dominerende sans hos de fleste fugle. To store optiske lapper ligger mellem hjernehalvdelene og lillehjernen, og det er her, at synsnerverne kommunikerer og formidler information. Der findes ikke noget corpus callosum, og hjernebarken er generelt meget tynd, men corpus striatum er veludviklet og menes at være stedet for mentale associationer hos fugle. Et vigtigt træk ved fuglehjernen er pinealkirtlen, som sidder i diencephalon, kranielt i forhold til lillehjernen i midterlinjen dorsalt. Pinealkirtlen har sekretoriske celler, der ligner fotoreceptorer, og den reagerer derfor på lys. De er også forbundet via de kraniale cervikale ganglier med synsnerven. Pinealkirtellegemet er ansvarlig for reguleringen af mange årstidsbestemte virkninger såsom reproduktion og migration samt cirkadiske rytmer. Det har en direkte virkning via hormonudskillelse på hypothalamus.
Figur 9.1 Dorsal side af fuglehjernes hjernehalvdele, der viser mangel på gyri. 
Det aviære nervesystem er ikke ulig det, der ses i dets modstykke hos pattedyr. Fugle besidder 12 kranienerver (CN), det samme antal som hos katte og hunde. Hos fugle er synsnerven (CN II) den største kranienerve, idet den er næsten halvt så stor som rygsøjlens diameter. Den passerer gennem kalvariet via et enkelt hul i stedet for gennem flere mindre huller i forbindelse med pattedyrenes cribriformplade. De øvrige 11 kranienerver har lignende funktioner som hos pattedyr, og læseren henvises til mere dybdegående anatomitekster for yderligere oplysninger (King & McLelland, 1975; Bennett, 1994).
Hver af vingerne har en nerveforsyning fra et brachialplexus, der stammer fra rygmarvsnerverne i det caudale cervikale område. Der findes tre nerveplexusser i lumbosakralregionen: lumbale, ischiatiske og pudendale. Det lumbale plexus stammer fra de to sidste lumbale og de to første sakrale spinalnerverødder. Ligesom de andre lumbosakrale plexusser ligger det i et hulrum i bækkenet, dorsalt for det kraniale nyreområde. Det forsyner kropsvæggen og de øvre benmuskler og giver anledning til obturator-, femoral-, kraniale gluteal- og saphenusnerve. I modsætning til hunde og katte har fugle et ischiatisk plexus, der stammer fra fire til syv rygmarvsnerver i sakralområdet, og som er placeret i et hulrum i bækkenet dorsalt for den midterste nyrestruktur. Det giver anledning til den vigtigste nerveforsyning af baglårene – iskiasnerven, som er den største perifere nerve i kroppen, og den caudale glutealnerve. Endelig dannes et plexus pudendal i et hulrum i bækkenet dorsalt for det kaudale nyreområde fra fem coccygeale spinalnerver og innerverer halen og cloacalområdet.
Muskuloskeletale system
De fleste fugle har flyveevne. De tætte, tunge knogler hos det jordbundne pattedyr ville kræve for stor kraftanstrengelse for at løfte sig op i luften. Fuglene har derfor tilpasset deres skeletstruktur, idet de har forenklet antallet af knogler ved at smelte nogle sammen og generelt lettet hele strukturen ved at skabe luftrum i mange af knoglerne.
For yderligere at lette skelettet er flere af de større knogler og endda nogle af ryghvirvlerne i rygsøjlen direkte eller indirekte forbundet med luftvejene og siges at være pneumoniserede. Dette erstatter det tykke marvhul eller knoglemarv, som findes i midten af pattedyrs knogler, og giver en let, trabekulær struktur. Selv om strukturen er let, er den ikke desto mindre ekstremt stærk.
Figur 9.2 viser et generaliseret fugleskelet.
Figur 9.2 Fugleskelet. 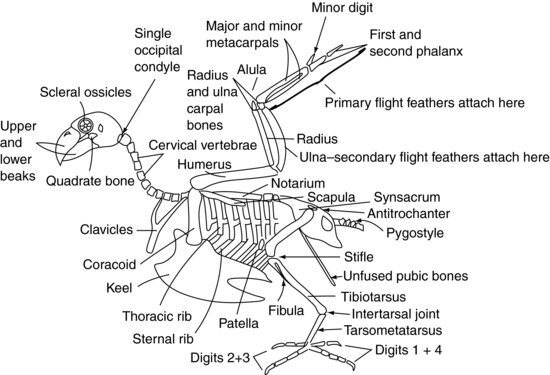
Skranie
Næb
Næbbet, eller næbbet, er det vigtigste kendetegn ved fugleskallen. Det er blevet modificeret i et forbløffende antal former og størrelser, hvilket hovedsageligt afhænger af den kost, som fuglen er blevet tilpasset til. I alle tilfælde består det af et over- (maxillary) og undernæb (mandibular), som er dækket af et lag keratin, en sej proteinforbindelse, der ligner den, som danner insekternes exoskelet. Dette keratinlag er kendt som rhamphotheca. Det er yderligere klassificeret således, at overkæbelaget kaldes rhinotheca, og underkæbelaget kaldes gnatotheca. Rhinotheca og gnatotheca vokser fra en plade ved basen af de respektive sider af næbbet, og udskiftningshastigheden afhænger af den type føde, der spises, og den slid, næbbet udsættes for.
I Psittaciformes (tabel 9.1) er overnæbbet kraftigt udviklet og ender i en skarp spids, der hænger ud over det bredere, kraftigere undernæb. Den enorme kraft i en papegøjes næb skyldes et synovialled eller en hængselsmekanisme, kendt som kinetisk led, som forbinder næbbet med kraniet. Papegøjens undernæb har en række tryksensorer i spidsen, som gør det muligt for den at teste konsistensen og strukturen af de genstande, den griber fat i.
Hos rovfugle er overnæbbet ekstremt skarpt og spidst, men mangler den kinetiske ledforbindelse, så den kan ikke producere en så kraftig nedadrettet kraft. I stedet bruges det som et riveinstrument.
I Anseriformes (andefamilien) er næbbet fladtrykt og kan have fine tandsnit i kanterne, som gør det muligt for fuglen at filtrere fine partikler fra vandet. Ænder som gråænder og skræppeænder har denne type næb. Disse tandsnit kan være videreudviklet til en savtakket kant (f.eks. hos den passende navngivne savsnæb-familie), som gør det muligt for fuglen at gribe glatte fødeemner, f.eks. fisk. Anseriformes har også nerveender i en plade i spidsen af næbbet (kendt som “neglen”), som gør det muligt for dem at finde føde, der er skjult i mudder.
I alle fugle er der en række mindre knogler bag det nederste og øverste næb, som gør det muligt for dem at bevæge næbbet uafhængigt af kraniet. Disse omfatter palatin-, quadrat- og pterygoid-knoglerne og jugalbuerne. Deres nøjagtige bevægelser ligger uden for denne tekst at beskrive, men mange af referencerne i slutningen af dette kapitel giver gode beskrivelser af deres funktion.
Næsebor
Næseborene eller næseborene ligger ved basen af det øverste næb hos de fleste fugle og er ofte omgivet af et område af fjerløs hud, der kaldes cere. Dette kan være stærkt farvet hos nogle arter, som f.eks. hos vipstjert, hvor de kan bruges til at identificere fuglens køn. Hos mange Anseriformes ligger næbbet mere mod næbbets spids. Selve næseborene er blot åbninger til bihulekamrene, som igen er forbundet med et forgrenet netværk af knoglekamre i hele fuglens hoved. Disse bihuler varierer alt efter art, men de fleste fuglepatienter har en infraorbital bihule. Denne sidder under øjnene og er ofte involveret i bihule- og øjeninfektioner. Den adskiller sig fra bihuler hos de fleste pattedyr ved, at den laterale væg ikke har nogen knogle, men kun er dækket af blødt væv. Dette betyder, at enhver infektion i infraorbital sinus ofte resulterer i en hævelse i fuglens ansigt ventral for øjet. Disse bihuler har også forbindelse med luftsække i hoved og hals. Funktionen af disse luftsække er ikke klarlagt, men de kan være med til at give stemmeresonans. Når en fugl lider af bihulebetændelse, kan de smalle indgange til disse bihuler blive delvist blokeret og fungere som envejsventiler, der tillader luft ind i bihullerne, men ikke ud. Sækkene kan derefter overoppustes, og der ses da ofte bløde hævelser på ryggen eller nakken af fuglens hoved.
Sinuserne og de ydre næsebor kommunikerer med oropharynx via choanalspalten. Dette er en smal åbning i den hårde ganes midterlinje og sidder umiddelbart over glottis, når næbbet er lukket, hvilket gør det muligt for fuglen at trække vejret gennem næseborene. Det er ofte det område, der vælges til at tage prøver, når man forsøger at isolere infektiøse agenser for sygdom i de øvre luftveje hos fugle.
Den fugles kranie er forbundet med atlas (eller den første ryghvirvel) via kun én baghovedkondyl ved kraniets basis, i modsætning til pattedyrenes to. Der er også et stort antal meget bevægelige halshvirvler. Disse to faktorer gør fuglehovedet meget bevægeligt. Imidlertid er det atlanto-occipitale led også et svagt punkt, hvilket gør dislokation på dette sted meget let.
Hvirvelsøjle
Halshvirvler
Halshvirvlerne (figur 9.2) er uafhængigt bevægelige hos fuglen, ligesom de er hos pattedyret, og varierer i antal alt efter art mellem 11 og 25. De er generelt kasseformede.
Torakal-, lænde- og sakralhvirvler
Torakalhvirvlerne (figur 9.2) er hos rovfugle, duer og mange andre arter smeltet sammen til en enkelt knogle, der kaldes notarium. Hos andre arter har de en vis begrænset bevægelighed. Der er så to ryghvirvelsamlinger mellem notarium og de sammenvoksede lænde- og korshvirvler. Disse sammenvoksede ryghvirvler er kendt som synsacrum. Synsacrum smelter sammen med selve bækkenet og danner et rygskjold af knogle over fuglens hale side.
Coccyghvirvler
De fleste af de caudale coccyghvirvler (Figur 9.2) er normalt smeltet sammen til en enkelt struktur kaldet pygostyle – som danner den “parsonsnæsede” del af kyllingen!
Bækken
Bækkenets tag udgøres af synsacrum (Figur 9.2). Bækkenets to “sider” er reduceret i størrelse i forhold til pattedyr, men består af hoftebenet og iskiasknoglerne, og acetabulum dannes der, hvor de mødes. Acetabulum hos fugle er ikke en komplet knoglehule som hos pattedyr, men en fibrøs plade. På den laterale side af bækkenet er der en kam, der er kendt som antitrochanter, som er forbundet med lårbensknoglens trochanter. Denne kam har til opgave at forhindre, at lemmerne abduceres, når man sidder på en stolpe. Bækkenets skamben smelter ikke sammen i den ventrale midterlinje, som det er tilfældet hos pattedyr. I stedet danner de fine lange knogler, som strækker sig kaudalt mod bugvæggen. De giver støtte til huden, der dækker det hale bagkrop, og giver plads nok til passage af æg hos hunfuglen.
Ribben
Pittaciformes har otte par ribben (Figur 9.2). Hvert ribben har et rygsegment, kendt som brystribben, og et ventralsegment, eller sternalribben. Disse ribben peger bagud og forbinder brysthvirvlerne stift dorsalt med kølen, eller sternum, ventralt.
Sternum
Sternumhvirvlerne er smeltet sammen hos fuglene for at danne kølen. Kølen har en midterste linjekam, som opdeler brystmusklerne i højre og venstre side. Ryggen kan være en dyb struktur, som det ses hos duer, rovfugle og Psittaciformes, hvilket gør det muligt for store brystmuskler at sætte sig fast for at sikre en stærk flyvning. Alternativt kan kølen være fladtrykt, som hos Anseriformes, for at give en bådlignende struktur, der er mere egnet til at flyde.
Vinger
Skulderleddet dannes af mødet mellem tre knogler: overarmsknoglen, skulderbladet (som er mere rørformet end det fladtrykte fra pattedyr) og en tredje knogle, der kaldes coracoid (Figur 9.2). Sidstnævnte knogle danner en støtte, som støtter skulderleddet mod brystbenet. Musculus supracoracoideus hæfter sig på kølen og passerer derefter gennem den foramen eller åbning, der er dannet ved disse knoglers mødested, og når således frem til overarmsknoglens dorsale side, hvor den hæfter sig. Kontraktion af denne muskel, sammen med nogle elastiske væv, som også er til stede, er med til at hæve vingen. Brystmusklerne sætter sig fast fra kølen på overarmsknoglen for at trække vingen nedad. De sammenvoksede kraveben eller ønskeben (ofte kaldet furcula) er forbundet med coracoidbenet og giver vingerne en vis grad af fjedervirkning i forbindelse med vingeslagene. Overarmsknoglen er pneumoniseret, hvilket betyder, at den ikke kan bruges til intraossøs væsketerapi. Dette er også et vigtigt punkt at overveje ved reparation af frakturer.
Humerus artikulerer med radius og ulna i albueleddet. Radius er den mindste af disse to knogler og ligger cranialt. Ulna udgør fastgørelseskilden for de sekundære flyvefjer, som indsættes direkte i periostet på denne knogle (Figur 9.3). Ulna bruges ofte til intraossøs væsketilførsel hos fugle.
Figur 9.3 Ventral side af en tårnfalkes (Falco tinnunculus) vinge med fjernede dækfjer, der viser de primære fjeres fastgørelse til hånden og de sekundære fjer til ulna. 
Radius og ulna artikulerer med henholdsvis en radial karpalkogle og en ulnar karpalkogle. Disse artikulerer igen med tre metacarpalknogler. Den første metacarpal knogle svarer til fuglenes “tommelfinger”. Den er kendt som alula eller “bastardvinge” og danner et fjerformet fremspring fra den kraniale side af carpometacarpalleddet. De resterende to metacarpalknogler er kendt som den store og den lille metacarpalknogle og er forbundet med det første fingerled kranielt og det lille fingerled kaudalt. Den første phalanx knyttes derefter sammen med den anden phalanx og danner vingespidsen. De primære fjer fæstner sig til periostet af phalanges og de mindre metacarpalknogler (Figur 9.3).
Vingens område er forstørret af tynde lag af elastisk væv, der spænder fra den ene ledflade til den anden. Det største strækker sig fra skulderen til karpalleddet kranialt og er kendt som propatagium eller “vingevæv” (Figur 9.4). Dette kan hos nogle arter, f.eks. duer, anvendes til administration af vaccine.
Figur 9.4 Dorsal side af en tårnfalkes (Falco tinnunculus) vinge med fjernede dækfjer, der viser det elastiske ark i propatagium, der bygger bro over albueleddet. 
Bækkenleddet
Bækkenets acetabulum holder lårbenshovedet (Figur 9.2). Lemmet kan låses og forhindres i at blive abduceret ved, at femurens trochanter major griber ind i den antitrochanteriske kam på bækkenet. Hos mange fugle er lårbenet pneumoniseret. I knæleddet er lårbenet i led med knæskallen og skinnebensknoglen. Tibiotarsalknoglen kaldes sådan, fordi den er dannet ved sammensmeltning af tibia og den proximale række af tarsalknogler, og den kan også bruges til intraossal væsketilførsel. På den laterale side af den proximale tibiotarsus sidder den meget reducerede fibula.
Distalt artikulerer tibiotarsus-knoglen med tarsometatarsus-knoglen. Denne knogle er dannet ved sammensmeltning af den distale række af tarsalknogler med den enlige metatarsalknogle. Leddet mellem tibiotarsus og tarsometatarsus er kendt som det intertarsale eller suffrago-led. Tarsometatarsus artikulerer derefter med phalanges.
I Psittaciformes peger to fingre fremad (det andet og tredje) og to bagud (det første og fjerde), hvilket skaber et zygodactyl lem. Det første finger har to phalanges, det andet finger har tre phalanges, det tredje finger har fire phalanges og det fjerde finger har fem phalanges. Hos flyvefugle (Passeriformes) og rovfugle peger andet, tredje og fjerde finger fremad, og det første finger peger bagud, hvilket giver et anisodactyl lem. Nogle arter, som f.eks. fiskeørn (Pandion haliaetus), kan flytte det fjerde finger til at pege fremad eller bagud for at hjælpe med at fange sit bytte, hvilket skaber et semi-zygodactyl lem.
Særlige sanser
Øje
Fugleøjet er unikt, idet det indeholder en række små knogler. Disse er kendt som de sclerale benknogler (Figur 9.2). De danner en ringformet struktur, som støtter øjets forreste del. Fugleøjet adskiller sig også fra pattedyrøjet ved, at det ikke er en globus, men pæreformet med den smallere ende yderst.
Fugleøjet er stort i forhold til kraniets samlede størrelse, og kun et papirtyndt knogleskillevæg adskiller højre og venstre øjenhule. Fuglene har et bevægeligt, gennemsigtigt tredje øjenlåg samt øvre og nedre øjenlåg, hvoraf det nedre er mere bevægeligt end det øvre. Der findes almindeligvis to tåreproducerende kirtler: det tredje øjenlåg eller Harderian-kirtlen, som er placeret ved basis af det tredje øjenlåg, og lacrimalkirtlen, der ligger caudolateralt, som hos pattedyr.
Farven på iris kan ændre farve med alderen hos nogle papegøjer, for eksempel har den afrikanske grå papegøje en mørkegrå iris indtil 4-5 måneders alderen, hvorefter den bliver gulgrå og derefter sølvfarvet, efterhånden som den bliver ældre. Hos andre kan iris bruges som en indikator for fuglens køn: hos store kakaduer har hunnen f.eks. en lys, rødbrun iris, mens hannens er mørk, brunsort.
Fuglenes nethinde er tyk og har ingen synlige overfladeblodkar, i modsætning til pattedyrenes nethinde. For at forsyne nethinden med næring har fuglene en plisseret og foldet karstruktur kaldet pecten oculi, som findes på det sted, hvor synsnerven går ind i øjet. Den trækker sig sammen med mellemrum og udstøder næringsstoffer i glaslegemet.
Endeligt har fuglenes iris skeletmuskelfibre i sig i modsætning til pattedyr, som kun har glatmuskelfibre. Dette betyder, at aviærpatienten kan indsnævre og udvide sin pupil efter behag, hvilket reducerer værdien af den pupillære lysrefleks som et redskab til at bestemme øjenfunktion. Da de to synsnerver er helt adskilt fra hinanden, er den konsensuelle lysrefleks også en dårlig indikator for hjernefunktionen.
Øre
Der findes ingen øresnegl hos fugle, selv om nogle arter, som f.eks. den lang- og korthårede ugle, har fjer i dette område. Der er en kort, vandret ydre kanal, der er dækket af fjer, og som er placeret caudolateralt i forhold til øjenhulen. Trommehinden kan ses tydeligt. Mellemøret er forbundet med oropharynx via den Eustachiske kanal. Pattedyrenes øreknogler er hos fuglen erstattet af en lateral, ekstra columella-brusk og en medial columella-knogle, som overfører lydbølger til det indre øre.
Det indre øre indeholder sneglen og de halvcirkulære kanaler, som har de samme funktioner som hos pattedyr.
Atmningsanatomi
Overste åndedrætsorganer
Narerne munder ud i næsepassagerne, som igen kommunikerer med strubehovedets glottis via en midterste åbning i den hårde gane, som danner taget af den kaudale mund. Denne åbning kaldes choanalspalten. Sinussystemet og de cervicocephale luftsække er tidligere blevet nævnt.
Larynx
Fugle har en reduceret strubehovedstruktur, idet de mangler en epiglottis, skjoldbruskkroppen og stemmelæberne, som ses hos katte og hunde. Den vigtigste struktur er glottis, som beskytter indgangen til luftrøret. Udvendige muskler trækker glottis og luftrøret fremad, så det kommunikerer direkte med choanalspalten, hvilket gør det muligt for fuglen at trække vejret gennem næseborene. Glottis holdes lukket i hvile og åbnes kun ved indånding og udånding.
Trachea
Fuglearternes trachea adskiller sig fra pattedyrenes trachea ved, at dens bruskringe er komplette, signetringformede cirkler, der griber ind i hinanden, i stedet for de C-formede ringe i pattedyrenes trachea. Hos Psittaciformes og dagaktive rovfugle er formen af disse bruskringe let fladtrykt i dorsoventral retning, mens de hos de fleste Passeriformes er runde.
I nogle arter, såsom sangsvane og perlehøne, danner luftrøret en række sløjfer og spiraler ved thorakalindløbet. Andre arter, som f.eks. emu’en, har en ventral deling af luftrøret i midterlinjen tre fjerdedele af afstanden mellem hovedet og thorakalindløbet. Trakealforingens slimhinde stikker ud gennem denne spalte og danner en trakealsæk. Dette forbedrer stemmens resonans. Hos hanænder, f.eks. gråænder, er der en hævelse i den sidste del af luftrøret, ofte lige inden for brystkassen, kendt som trakealbulla.
Syrinx
For luftrøret deler sig i de to hovedbronkier, er der en struktur kendt som syrinx (Figur 9.6). Det er her, at fuglen producerer det meste af sin stemme. Den består af en række muskler og to membraner, som kan vibreres, uafhængigt af indånding eller udånding.
Nedre åndedrætsorganer
Lunger
Lungerne hos fuglearter er stive i deres struktur og puster sig ikke op eller puster sig ikke væsentligt op eller ned. De er fladtrykte i form og er fast knyttet til den ventrale side af brysthvirvlerne og hvirvelribberne. Der er ikke noget mellemgulv hos fugle, og den fælles kropshule kaldes coelom.
De parrede bronkier er støttet af C-formede ringe af brusk, i modsætning til luftrøret. De primære bronkier forsyner hver af de to lunger og deler sig hurtigt i sekundære og tertiære bronkier, eller parabronchi. Der er fire hovedgrupper af sekundære bronkier, der forsyner lungerne, men deres rolle i gasudvekslingen er minimal. De tertiære bronkier spiller dog en rolle i gasudvekslingen, da deres vægge er fyldt med membraner, der er i stand til at udveksle gasser. Disse områder fremstår som små huller, eller forkamre, hvortil der er forbundet endnu finere rør, der kaldes luftkapillærer. Disse er sammenflettet med hinanden og danner et tredimensionelt net, der er sammenvævet med blodkapillærbedene. Disse luftkapillærer varierer i størrelse, men har i gennemsnit en diameter på ca. 3-5 mm. Denne ekstremt lille diameter giver meget store tiltrækningskræfter mellem deres vægge, når der er flydende sekreter til stede, hvilket resulterer i hurtig blokering af luftvejsoverfladerne. For at forhindre, at dette sker, er der celler i parabronchi, som udskiller surfaktant for at sikre, at luftvejene forbliver åbne.
Lungestrukturen kan yderligere inddeles efter luftstrømmens retning i den i den neopulmoniske lunge og den paleopulmoniske lunge. Disse vil blive nævnt senere, når vi diskuterer respiratorisk fysiologi.
Luftsække
Den sidste del af fuglenes nedre åndedrætsorganer består af luftsækkene (se figur 9.5). Disse er ballonlignende sække, der fungerer som bælge og pumper luften ind og ud af de stive fuglelunger som reaktion på bevægelser af kropsvæggen og brystbenet. Luftsækkens vægge er meget tynde og består af simpelt pladeepithel, som dækker et lag dårligt vaskulariseret elastisk bindevæv.
Figur 9.5 Fuglens luftsækkesystem hos en and: (a) næsepassager, (b) infra-orbital sinus, (c) cervicocephale luftsække (enkelt), (d) claviculære luftsække, (e) kraniale thorakale luftsække, (f) kaudale thorakale luftsække og (g) abdominale luftsække. H, hjerte; L, lever; Lu, lunger; B, sprøjtebølle (hanænder).
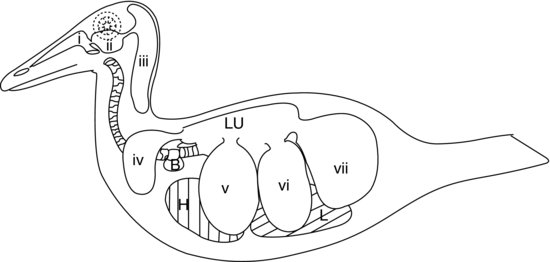
I de fleste fugle er der ni luftsække. En af disse er den allerede nævnte separate luftsæk, den cervicocephale luftsæk, som slet ikke kommunikerer med lungerne. De otte andre luftsække kommunikerer alle med lungerne via en sekundær bronchus (undtagen de abdominale luftsække, som har forbindelse til den primære bronchus på hver side). Figur 9.5 viser luftsækkesystemet hos en and.
Ud over den separate cervicocephale luftsæk er de andre otte standardluftsække