Veterian Key
- Familjen Anatidae, underfamiljen Anatinae, t.ex. gräsand, späckhuggare, ejderänder och shelducks
- Familjen Anatidae, underfamiljen Anserinae, Tribe Anserini (svanar och sädgäss), till exempel stumma svanar, sångsvanar och Bewickssvanar samt grågäss
- Falconidae-familjen till exempel pilgrimsfalk, saker, llanner, jaktfalken
- Familjen Accipitridae, t.ex. gamar (vanlig, grovben och honung), sparvhök, goshawk, kungsörn
Psittaciformes är bland de mest färgglada fåglarna som hålls som sällskapsdjur.
Nervsystemet
Fågelns hjärna är extremt slät och saknar de många gyri (hjärnans åsar) som ses hos däggdjur (figur 9.1). Synen verkar vara det dominerande sinnet hos de flesta fåglar. Två stora optiska lober ligger mellan hjärnhalvorna och lillhjärnan, och det är här som de optiska nerverna kommunicerar och sprider information. Det finns inget corpus callosum, hjärnbarken är i allmänhet mycket tunn, men corpus striatum är välutvecklat och tros vara platsen för mentala associationer hos fåglar. Ett viktigt inslag i fågelhjärnan är pinealkroppen som sitter i diencephalon, kranial till lillhjärnan i mitten av ryggraden. Tallkroppen har sekretoriska celler som liknar fotoreceptorer och reagerar därför på ljus. De är också kopplade via de kraniala cervikala ganglierna till synnerven. Pinealkroppen är ansvarig för regleringen av många säsongsbetonade effekter som reproduktion och migration samt cirkadiska rytmer. Den har en direkt effekt via hormonutsöndring på hypotalamus.
Figur 9.1 Dorsal aspekt av fågelns hjärnhalvor som visar avsaknad av gyri.

Det fågelns nervsystem är inte olikt det som ses hos dess motsvarighet hos däggdjur. Fåglar har 12 kranialnerver (CN), samma antal som hos katter och hundar. Hos fåglar är synnerven (CN II) den största kranialnerven, med en diameter som är nästan hälften av ryggraden. Den passerar genom kalvariet via ett enda hål i stället för genom flera mindre hål i samband med den kribriformiska plattan hos däggdjur. De övriga 11 kranialnerverna har liknande funktioner som hos däggdjur, och läsaren hänvisas till mer djupgående anatomitexter för mer information (King & McLelland, 1975; Bennett, 1994).
Varje vinge har nervförsörjning från ett plexus brachialis som härstammar från ryggmärgsnerverna i det kaudala cervikala området. Det finns tre nervplexus i den lumbosakrala regionen: lumbala, ischiatiska och pudendala. Plexus lumbalis härstammar från de två sista lumbala nervrötterna och de ett till två första sakrala nervrötterna. Liksom de andra lumbosakrala plexusarna ligger den i en hålighet i bäckenet, dorsalt i förhållande till det kraniala njurområdet. Det försörjer kroppsväggen och musklerna i övre benet och ger upphov till nerverna obturator, femoral, kranial gluteal och saphena. Till skillnad från hundar och katter har fåglar ett ischiatiskt plexus som härstammar från fyra till sju spinalnerver i sakralområdet och som ligger i en hålighet i bäckenet dorsalt i förhållande till den mellersta njurstrukturen. Det ger upphov till den huvudsakliga nervförsörjningen för bakbenen – ischiatnerven, som är den största perifera nerverna i kroppen, och den kaudala glutealnerven. Slutligen bildas ett plexus pudendal i en hålighet i bäckenet dorsalt för det kaudala njurområdet av fem spinalnerver från coccygealspinalnerver och innerverar svansen och cloacalområdet.
Muskuloskeletala systemet
De flesta fåglar har flygförmåga. De jordbundna däggdjurens täta och otympliga ben skulle kräva för mycket ansträngning för att lyftas upp i luften. Fåglar har därför anpassat sin skelettstruktur, förenklat antalet ben genom att smälta samman vissa och generellt sett lätta hela strukturen genom att skapa luftutrymmen inom många av benen.
För att ytterligare lätta skelettet är flera av de större benen, och till och med en del av ryggkotorna i ryggraden, direkt eller indirekt kopplade till luftvägarna, och sägs vara pneumoniserade. Detta ersätter det tjocka märghålan eller benmärgen som finns i mitten av däggdjursbenen och ger en lätt, trabekulär struktur. Även om strukturen är lätt är den ändå extremt stark.
Figur 9.2 visar ett generaliserat skelett av fåglar.
Figur 9.2 Fågelskelett.
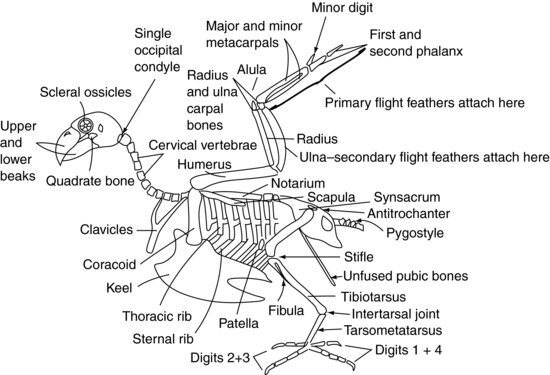
Skalle
Näbb
Näbben, eller näbben, är det främsta kännetecknet på fågelskallen. Den har modifierats till ett förbryllande antal former och storlekar, främst beroende på den diet som fågeln har anpassat sig till. I alla fall består den av en övre (maxillär) och en undre (mandibulär) näbb som är täckta av ett lager keratin, en seg proteinförening som liknar den som bildar insekternas exoskelett. Detta keratinskikt kallas rhamphotheca. Det klassificeras vidare så att maxillära lagret kallas rhinoteka och mandibulära lagret gnatoteka. Rhinotheca och gnatotheca växer från en platta vid basen av de respektive sidorna av näbben, och utbyteshastigheten beror på vilken typ av föda som äts och den nötning som näbben utsätts för.
I Psittaciformes (tabell 9.1) är övernäbben kraftfullt utvecklad och slutar i en skarp spets som överskjuter den bredare, kraftigare undernäbben. Den enorma kraften i papegojans näbb beror på en synovialled eller gångjärnsmekanism, den så kallade kinetiska leden, som förenar näbben med skallen. Papegojans nedre näbb har en rad trycksensorer i sin spets, vilket gör att den kan testa konsistensen och strukturen hos de föremål den tar tag i.
Hos rovfåglar är den övre näbben extremt vass och spetsig, men den saknar den kinetiska ledanslutningen och kan därför inte åstadkomma en lika kraftfull nedåtgående kraft. Istället används den som ett rivningsinstrument.
I Anseriformes (ankfamiljen) är näbben tillplattad och kan ha fina tandningar i kanterna som gör det möjligt för fågeln att filtrera fina partiklar från vattnet. Ankor som gräsänder och skator har den här typen av näbb. Dessa tandningar kan vidareutvecklas till en tandad kant (t.ex. hos familjen med det passande namnet sågskallra) som gör det möjligt för fågeln att ta tag i hal mat, t.ex. fisk. Anseriformes har också nervändar i en platta vid näbbens spets (känd som ”spiken”) som gör det möjligt för dem att hitta föda som är gömd i lera.
I alla fåglar finns det en serie mindre ben bakom den nedre och övre näbben som gör det möjligt för dem att röra näbben oberoende av skallen. Dessa inkluderar palatin-, quadrate- och pterygoidbenen samt jugalbågarna. Deras exakta rörelser ligger utanför denna text att beskriva, men många av referenserna i slutet av detta kapitel ger bra beskrivningar av deras funktion.
Näsborrar
Näsborrarna, eller näsborrarna, ligger vid basen av övernäbben hos de flesta fåglar och är ofta omgivna av ett område med fjäderlös hud som kallas cere. Detta kan vara starkt färgat hos vissa arter, t.ex. hos gräshoppan, där de kan användas för att identifiera fågelns kön. Hos många Anseriformes ligger näsan mer mot näbbens spets. Näsborrarna i sig är bara öppningar till sinuskamrarna, som i sin tur ansluter till ett förgrenat nätverk av benkamrar i hela fågelhuvudet. Dessa bihålor varierar beroende på art, men majoriteten av fågelpatienterna har en infraorbital bihåla. Denna sitter under ögonen och är ofta inblandad i bihåleinflammationer och ögoninfektioner. Den skiljer sig från bihålorna hos de flesta däggdjur genom att sidoväggen inte har något ben utan endast är täckt av mjuk vävnad. Detta innebär att en infektion i infraorbitala bihålor ofta resulterar i svullnad på fågelns ansikte ventralt om ögat. Dessa bihålor har också kontakt med luftsäckar i huvudet och halsen. Funktionen hos dessa luftsäckar är oklar, men de kan hjälpa till med röstresonansen. När en fågel lider av bihåleinflammationer kan de smala inloppen till dessa bihålor bli delvis blockerade och fungera som enkelriktade ventiler som släpper in luft i bihålorna men inte ut. Säckarna kan då överblåsas, och mjuka svullnader ses då ofta på baksidan eller nacken av fågelns huvud.
Sinusarna och de yttre näsorna står i förbindelse med oropharynx via choanalspalten. Detta är en smal öppning i mitten av den hårda gommen och sitter omedelbart över glottis när näbben är stängd, vilket gör att fågeln kan andas genom näsborrarna. Det är ofta det område som väljs för att ta prover när man försöker isolera smittämnen för sjukdomar i de övre luftvägarna hos fåglar.
Den aviära patientens skalle är ansluten till atlasen (eller den första ryggkotan) via endast en occipitalkondyl vid skallbasen, till skillnad från däggdjurens två. Det finns också ett stort antal mycket rörliga halsryggkotor. Dessa två faktorer gör fågelhuvudet extremt rörligt. Men atlanto-occipitalleden är också en svag punkt, vilket gör att dislokation på den platsen är mycket lätt.
Verbralpelaren
Halsryggkotorna
Halsryggkotorna (figur 9.2) är självständigt rörliga hos fågeln, precis som hos däggdjuren, och varierar beroende på art mellan 11 och 25 i antal. De har i allmänhet en boxliknande form.
Brygg-, länd- och sakralkotor
Bryggkotorerna (figur 9.2) är sammanfogade hos rovfåglar, duvor och många andra arter för att bilda ett enda ben som kallas notarium. Hos andra arter har de en viss begränsad rörlighet. Det finns sedan två ryggmärgsfogar mellan notarium och de sammanfogade länd- och sakralkotorna. Dessa sammanfogade ryggkotor kallas synsacrum. Synsacrum smälter samman med själva bäckenet och bildar en dorsal sköld av ben över den kaudala sidan av fågeln.
Coccygealkotorna
Majoriteten av de kaudala coccygealkotorna (Figur 9.2) är vanligtvis sammanfogade till en enda struktur som kallas pygostyle – som bildar den ”parsonsnäsiga” delen av kycklingen!
Bäckenet
Bäckenets tak bildas av synsacrum (figur 9.2). Bäckenets två ”sidor” är mindre i storlek jämfört med däggdjur men består av iliakal- och ischialbenen, och acetabulum bildas där de möts. Acetabulum hos fåglar är inte en fullständig benhåla som hos däggdjur, utan ett fibröst skikt. På bäckenets sida finns det en kam som kallas antitrochanter, som är i led med lårbenets större trochanter. Denna kam har till uppgift att förhindra att extremiteten abduceras när man sitter på en stolpe. Bäckenets blygdben smälter inte samman i den ventrala mittlinjen som hos däggdjur. I stället bildar de fina långa ben som sträcker sig kaudalt mot ventrikeln. De ger stöd åt huden som täcker den kaudala buken och tillräckligt med utrymme för passage av ägg hos honfågeln.
Ribkorg
Psittaciformes har åtta par revben (figur 9.2). Varje revben har ett dorsalt segment som kallas bröstribban och ett ventralt segment, eller sternalribban. Dessa revben pekar bakåt och förbinder bröstkotorna styvt dorsalt och kölen, eller bröstbenet, ventralt.
Sternum
Sternumkotorna smälter samman hos fåglar för att bilda kölen. Kielet har en mittlinjenrygg som delar upp bröstmusklerna i höger och vänster sida. Ryggen kan vara en djup struktur, vilket ses hos duvor, rovfåglar och Psittaciformes, vilket gör det möjligt för stora bröstmuskler att fästa för stark flygning. Alternativt kan kölen vara tillplattad, som hos Anseriformes, för att ge en båtliknande struktur som är mer lämpad för att flyta.
Vingar
Skulderleden bildas av mötet mellan tre ben: överarmsbenet (humerus), skulderbladet (som är mer rörformigt än det tillplattade däggdjursbenet) och ett tredje ben, det s.k. koracoidbenet (figur 9.2). Det sistnämnda benet bildar en stomme som stödjer axelleden mot bröstbenet. Musculus supracoracoideus fäster vid kölen, passerar sedan genom den foramen, eller öppning, som bildas vid mötespunkten för dessa ben och når på så sätt överarmsbenets dorsala sida där den fäster. Kontraktion av denna muskel, tillsammans med vissa elastiska vävnader som också är närvarande, hjälper till att höja vingen. Bröstmusklerna fäster från kölen på överarmsbenet för att dra vingen nedåt. De sammanfogade nyckelbenen, eller önskebenet (ofta kallat furcula), är sammanfogade med korakoidbenet och ger vingarna en viss fjädring när de flaxar med vingarna. Överarmsbenet är pneumoniserat, vilket innebär att det inte kan användas för intraosseös vätskebehandling. Detta är också en viktig punkt att ta hänsyn till vid reparation av frakturer.
Humerus artikulerar med radius och ulna i armbågsleden. Radius är det mindre av dessa två ben och ligger cranialt. Ulna utgör fästkällan för de sekundära flygfjädrarna, som sätter sig direkt i detta benets periosteum (figur 9.3). Ulna används ofta för intraosseös vätskeadministrering hos fåglar.
Figur 9.3 Ventral aspekt av en tornfalks (Falco tinnunculus) vinge med täckfjädrar avlägsnade som visar primärfjädrarnas infästning i manus och sekundärfjädrarna i ulna.

Radius och ulna artikulerar med ett radialt karpalben respektive ett ulnar karpalben. Dessa artikulerar i sin tur med tre metakarpalben. Det första metakarpalbenet är motsvarigheten till fågelns ”tumme”. Det är känt som alula, eller ”bastardvinge”, och bildar en fjäderformad utskjutning från den kraniala sidan av carpometacarpalleden. De återstående två metacarpalbenen kallas stora och lilla metacarpalbenen och är knutna till det första falangen kranialt och det lilla fingret kaudalt. Den första falangen artikulerar sedan med den andra falangen och bildar vingspetsen. De primära fjädrarna fäster vid periostet av phalangerna och de mindre metakarpalbenen (figur 9.3).
Vingens yta förstoras av tunna skikt av elastisk vävnad som sträcker sig från den ena ledytan till den andra. Det största sträcker sig från axeln till karpalleden kraniellt och kallas propatagium eller ”vingväv” (figur 9.4). Detta kan användas hos vissa arter, till exempel duvor, för administrering av vaccin.
Figur 9.4 Dorsal sida av en tornfalks (Falco tinnunculus) vinge med täckfjädrar borttagna som visar det elastiska bladet i propatagium som överbryggar armbågsleden. 
Bäckenled
Bäckenets acetabulum rymmer lårbenshuvudet (figur 9.2). Extremiteten kan låsas och hindras från att abduceras genom att lårbenets större trokanter griper in i den antitrokantära kammen på bäckenet. Lårbenet är pneumoniserat hos många fåglar. I knäleden är lårbenet i led med knäskålen och skenbenet. Tibiotarsalbenet kallas så eftersom det bildas genom sammansmältning av tibia och den proximala raden av tarsalben, och kan också användas för intraosseös vätskeadministrering. På den laterala sidan av det proximala tibiotarsusbenet finns den mycket reducerade fibula.
Distalt artikulerar tibiotarsusbenet med tarsometatarsusbenet. Detta ben bildas genom sammansmältning av den distala raden av tarsalben med det ensamma metatarsalbenet. Leden mellan tibiotarsus och tarsometatarsus är känd som intertarsalleden eller suffragoleden. Tarsometatarsus artikulerar sedan med phalanges.
I Psittaciformes pekar två fingrar framåt (den andra och tredje) och två bakåt (den första och fjärde), vilket skapar en zygodactyl lem. Den första fingern har två falanger, den andra fingern har tre falanger, den tredje har fyra falanger och den fjärde har fem falanger. Hos fåglar som flyger (Passeriformes) och rovfåglar pekar den andra, tredje och fjärde fingret framåt och den första bakåt, vilket ger en anisodactyl lem. Vissa arter, till exempel fiskgjuse (Pandion haliaetus), kan flytta den fjärde fingret så att det pekar framåt eller bakåt för att underlätta fångsten av bytet, vilket skapar en semi-zygodactyl lem.
Speciella sinnen
Öga
Fågelns öga är unikt genom att det innehåller en rad små ben. Dessa är kända som de sklerala benstommarna (figur 9.2). De bildar en ringformad struktur som stöder ögats framsida. Fågelögat skiljer sig också från däggdjursögat genom att det inte är en glob utan päronformat, med den smalare änden längst ut.
Fågelögat är stort i förhållande till skallens totala storlek, med endast en papperstunn benig skiljevägg som skiljer den högra och vänstra ögonhålan åt. Fåglar har ett rörligt, genomskinligt tredje ögonlock samt övre och nedre ögonlock, varav det nedre är mer rörligt än det övre. Två tårproducerande körtlar finns vanligen: det tredje ögonlocket, eller Harderianska körteln, som ligger vid basen av det tredje ögonlocket, och tårkörteln som ligger kaudolateralt, som hos däggdjur.
Färgen på iris kan förändras med åldern hos vissa papegojor, t.ex. har den afrikanska grå papegojan en mörkgrå iris fram till 4-5 månaders ålder, då den blir gulgrå, och sedan silverfärgad när den fortsätter att åldras. Hos andra kan iris användas som en indikator på fågelns kön: hos stora kakaduor, till exempel, har honan en ljus, rödbrun iris, medan hanens är mörk, brunsvart.
Fågelns näthinna är tjock och har inga synliga ytliga blodkärl, till skillnad från däggdjurens näthinna. För att förse näthinnan med näring har fåglar en veckad och veckad kärlstruktur som kallas pecten oculi, som finns vid den punkt där synnerven går in i ögat. Den drar ihop sig med jämna mellanrum och stöter ut näringsämnen i glaskroppen.
För det sista har fåglars iris skelettmuskelfibrer i sig, till skillnad från däggdjur som endast har glatta muskelfibrer. Detta innebär att den aviära patienten kan dra ihop och utvidga sin pupill efter behag, vilket minskar värdet av den pupillära ljusreflexen som ett verktyg för att fastställa okulär funktion. Eftersom de två optiska nerverna är helt separerade från varandra är den konsensusliknande ljusreflexen också en dålig indikator på hjärnans funktion.
Öra
Det finns ingen pinna hos fåglar, även om vissa arter, t.ex. den långörade och den kortörade ugglan, har fjädrar i detta område. Det finns en kort, horisontell yttre kanal, täckt av fjädrar, som är belägen caudolateralt i förhållande till ögonhålan. Träthinnan kan ses tydligt. Mellanörat ansluter till orofarynx via den Eustachiska kanalen. Däggdjurens hörselhörnstenar ersätts hos fågeln av ett lateralt, extra columellabrosk och ett medialis columellaben som överför ljudvågor till innerörat.
Innerörat innehåller cochlea och de halvcirkulära kanalerna, som fyller samma funktioner som hos däggdjur.
Aandningsanatomi
Övre andningsorganen
Näsborrarna mynnar ut i näsgångarna, som i sin tur står i förbindelse med struphuvudets glottis via en öppning i mitten av linjen i den hårda gommen som utgör taket på den kaudala munnen. Denna öppning kallas choanalspalten. Sinussystemet och de cervicocefala luftsäckarna har nämnts tidigare.
Larynx
Fåglar har en reducerad struphuvudstruktur och saknar epiglottis, sköldkörtelns broskbrosk och de stämband som ses hos katter och hundar. Den viktigaste strukturen är glottis, som skyddar ingången till luftstrupen. Yttre muskler drar glottis och luftstrupen framåt så att den kommunicerar direkt med chokalspalten, vilket gör att fågeln kan andas genom näsborrarna. Glottis hålls stängd i vila och öppnas endast vid inandning och utandning.
Trakea
Fågelarternas trakea skiljer sig från däggdjurens trakea genom att broskringarna är kompletta, signetringformade cirklar, som griper in i varandra, i stället för de C-formade ringarna i däggdjurens trakea. Hos Psittaciformes och dagaktiva rovfåglar är formen på dessa broskringar något tillplattad i dorsoventral riktning, medan de hos de flesta Passeriformes är runda.
I vissa arter, som t.ex. sångsvanen och pärlhönan, bildar luftstrupen en serie slingor och spiraler vid bröstkorgsintaget. Andra arter, till exempel emu, har en ventral delning av luftstrupen i mitten av linjen tre fjärdedelar av avståndet mellan huvudet och bröstintaget. Trakealens slemhinna sticker ut genom denna spricka för att bilda en trakealsäck. Detta förbättrar röstresonansen. Hos hanar av ankor, t.ex. gräsänder, finns det en svullnad i den sista delen av luftstrupen, ofta precis innanför bröstkorgen, som kallas trakealbulla.
Syrinx
För att luftstrupen ska dela sig i de två huvudbronscherna finns det en struktur som kallas syrinx (figur 9.6). Det är här som fågeln producerar det mesta av sin röst. Den består av en serie muskler och två membran som kan vibreras, oberoende av inandning eller utandning.
Nedre andningsorgan
Lungor
Lungorna hos fågelarter är stela i sin struktur och blåser inte upp eller släpper luft nämnvärt. De är tillplattade till formen och sitter fast på den ventrala sidan av bröstkotorna och ryggkotornas revben. Det finns inget diafragma hos fåglar, och den gemensamma kroppshålan kallas coelom.
De parade bronkerna stöds av C-formade ringar av brosk, till skillnad från luftstrupen. De primära bronkerna försörjer var och en av de två lungorna och delar sig snabbt i sekundära och tertiära bronker, eller parabronker. Det finns fyra huvudgrupper av sekundära bronkier som försörjer lungorna, men deras roll i gasutbytet är minimal. De tertiära bronkierna spelar dock en roll i gasutbytet eftersom deras väggar är fyllda med membran som kan utbyta gaser. Dessa områden ser ut som små gropar, eller atria, till vilka är anslutna ännu finare rör som kallas luftkapillärer. Dessa flätar ihop sig med varandra och bildar ett tredimensionellt nät som är sammanvävt med blodets kapillärbäddar. Dessa luftkapillärer varierar i storlek men har en genomsnittlig diameter på 3-5 mm. Den extremt lilla diametern ger upphov till mycket höga attraktionskrafter mellan deras väggar när det finns vätskesekret, vilket resulterar i en snabb blockering av andningsytorna. För att förhindra att detta inträffar finns det celler i parabronchi som utsöndrar surfaktant för att se till att luftvägarna hålls öppna.
Lungestrukturen kan vidare klassificeras efter luftflödets riktning inom den i neopulmonisk lunga och paleopulmonisk lunga. Dessa kommer att nämnas senare när vi diskuterar andningsfysiologi.
Luftsäckar
Den sista delen av fåglarnas nedre andningsorgan består av luftsäckarna (se figur 9.5). Dessa är ballongliknande säckar som fungerar som bälgar och pumpar luften in och ut ur de stela fågellungorna som svar på rörelser av kroppsväggen och bröstbenet. Luftsäckarnas väggar är mycket tunna och består av enkelt skivepitel som täcker ett lager av dåligt vaskulariserad elastisk bindväv.
Figur 9.5 Aviärt luftsäcksystem hos en anka: a) näsgångar, b) sinus infra-orbitalis, c) cervicocefala luftsäckar (enstaka), d) klavikulära luftsäckar, e) kraniala thorakala luftsäckar, f) kaudala thorakala luftsäckar och g) abdominala luftsäckar. H, hjärta; L, lever; Lu, lungor; B, sprutbulla (hanar av ankor).
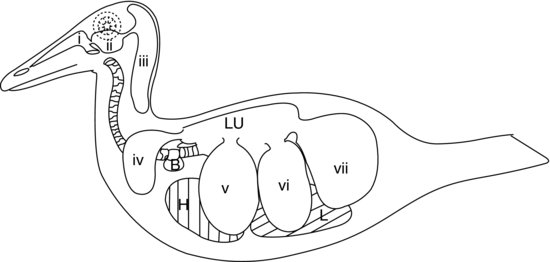
I de flesta fåglar finns det nio luftsäckar. En av dessa är den separata luftsäck som redan nämnts, den cervicocefala luftsäcken, som inte alls kommunicerar med lungorna. De övriga åtta kommunicerar alla med lungorna via en sekundär bronk (med undantag för bukluftsäckarna som ansluter till den primära bronken på varje sida). Figur 9.5 visar luftsäckssystemet hos en anka.
Förutom den separata cervicocefala luftsäcken är de övriga åtta standardluftsäckarna
.