Frontiers in Systems Neuroscience
Verschuivingen in de hersentoestand zijn geassocieerd met microcircuitveranderingen in neuronale vuren en met veranderingen op macroniveau in synchrone of asynchrone patronen van hersenactivatie. In dit kader zijn axonen relatief minder onderzocht (Barry, 2015), en dan voornamelijk in de context van geleidingssnelheid van actiepotentialen en de verhoogde alertheid die het gevolg kan zijn van veranderingen in geleidingssnelheid (e.g., Stoelzel et al., 2017). Daarnaast lijkt de sterk vertakte topologie van veel axonen echter goed ontworpen om gedistribueerde postsynaptische neuronale groepen differentieel te rekruteren, mogelijk op een toestandsafhankelijke manier. Signaalverwerking kan plaatsvinden in ten minste drie axonale domeinen; namelijk mapping, amplification, en timing (Innocenti et al., 2016; Innocenti, 2017). In dit Perspectief bespreek ik eerst kort netwerkvoorbeelden van axon collateralisatie, en vervolgens enkele van de axon-intrinsieke kenmerken die ten grondslag zouden kunnen liggen aan differentiële postsynaptische rekrutering. Omdat gedetailleerde parameters of mechanismen ontbreken, is het doel vooral om algemene kenmerken te belichten die een rol zouden kunnen spelen bij de controle en overgangen van hersentoestanden.
Axoncollateralen
Alle axonen hebben een uitgebreide distale arborisatie in de doelstructuur. Veel axonen hebben bovendien meerdere vertakkingen (ook wel collateralen genoemd) die gericht zijn op duidelijk verschillende structuren (besproken in Rockland, 2013, 2018). Een goed voorbeeld is de vertakking van axonen van laag 5 corticothalamus neuronen. Dit is herhaaldelijk gedocumenteerd en wordt vaak besproken als een belangrijk mechanisme waarmee een organisme kan onderscheiden of veranderingen in zintuiglijke input worden geproduceerd door veranderingen in de omgeving of door zelf geïnitieerde bewegingen (“efference copy” of “collateral discharge,” Guillery en Sherman, 2011).
In knaagdieren, waar er een groot aantal toepasbare technieken is om deze kwestie te onderzoeken, is bekend dat collateralisatie relatief vaak voorkomt. Om enkele voorbeelden te geven: (1) neuronen in het hippocampus subiculum projecteren naar de mammillaire lichaampjes, de retrospleniale cortex, of door collateralen naar beide (rat: Kinnavane et al, 2018); (2) serotonerge neuronen in de dorsale raphe (DR) kern maken in verschillende combinaties contact met het striatum, de prefrontale cortex, en de amygdala (rat: Gagnon en Parent, 2014), evenals met combinaties van autonome kernen (Waselus et al., 2011). In dit geval zou activering van de stressrespons door DR collateralen kunnen leiden tot gesynchroniseerde activatie van kernen die geassocieerd zijn met neurohormoonafgifte of pressorresponsen. Differentiële, gecoördineerde activeringen van voorhersenstructuren kunnen bijdragen aan de veelzijdige maar verwante DR-functies, zoals regulatie van de slaap-waakcyclus, modulatie van pijnsignalen, of stemmingsuitdrukking (Gagnon en Parent, 2014).
Derde, anatomische studies van thalamocorticale projecties bij muizen identificeren “multispecifieke axonen” die zich wijd vertakken naar beperkte domeinen in afzonderlijke corticale (en subcorticale) gebieden. Deze zijn verondersteld om de snelle opkomst en herconfiguratie van ruimtelijk gedistribueerde, synchroniseerbare neurale assemblages te orkestreren (Clasca et al., 2016).
Vierde, een recente studie van corticocorticale verbindingen met behulp van whole-brain axonal tracing in muis visuele cortex vond dat 23 van 30 neuronen contact maakten van twee tot zeven andere corticale gebieden. Ter bevestiging van dit resultaat vond high-throughput DNA-sequencing van genetisch gebarcodeerde neuronen dat 44% van 533 neuronen meervoudig projecterend waren (“broadcast neurons,” Han et al., 2018). Han et al. onderscheidden voorlopig twee brede types van projecterende neuronen, een kleinere “toegewijde” (uni-target) subpopulatie, naast een prevalentie van “omroepende” (meervoudig projecterende) cellen. Zou deze architectuur modulaties in cognitieve toestand en zintuiglijke verwerking kunnen ondersteunen?
Niet-gestereotypeerde collateralisatie
Zoals hierboven opgemerkt, neuronen die collateraliseren doen dit in een niet-stereotype patroon. Binnen een aangewezen projectie (gedefinieerd door de oorsprong), zenden neuronen takken naar een gediversifieerde subset van doelgebieden (“in alle combinaties”). Deze waarneming is zo consistent dat zij als een regel kan worden beschouwd, hoewel de gedetailleerde parameters niet in een tabel zijn opgenomen. De functionele betekenis is ook onduidelijk, maar opkomende resultaten suggereren dat de heterogene en variabele inputs naar een corticaal neuron (en, bij uitbreiding, zouden we kunnen afleiden naar neurale assemblages) belangrijk zijn in het aansturen van variabiliteit en spike trein veranderingen in de loop van experimentele trials (Gomez-Laberge et al, 2016 en zie hieronder).
Intrinsieke collateralen
In alle soorten, lange afstand corticale projectie neuronen, naast enkele of meerdere extrinsieke doelen, hebben meestal een uitgebreide intrinsieke arborization. Sommige corticale neuronen hebben alleen een intrinsieke (lokale) collateralisatie, en sommige alleen een extrinsieke (rat: Kita en Kita, 2012). Hoewel er slechts schaarse gegevens zijn voor de werkelijke verhoudingen van intrinsieke en extrinsieke arborizaties (Parent et al., 2000; Rockland, 2018), suggereert bewijs dat dit zeer variabel zal zijn. Zelfs binnen het systeem van intrinsieke verbindingen zijn er niet alleen verschillen in aantal collateralen en aantal synaptische boutons, maar een enkel neuron kan een mix van gemyeliniseerde en ongemyeliniseerde collateralen hebben (kat visuele cortex: Martin et al., 2014; Koestinger et al., 2017). Tak-specifieke myelinisatie zou kunnen resulteren in verhoogde, tak-specifieke geleidingssnelheid, hoewel Koestinger et al. suggereren dat het meer te maken kan hebben met factoren zoals verhoogde zekerheid van transmissie, vermoedelijk, wederom, tak-specifiek.
Een merkwaardige observatie met betrekking tot myelinisatie betreft de stria van Gennari, de gemyeliniseerde band van axonen in laag 4B van primatengebied V1. Aangezien deze bestaat uit intrinsieke collateralen, is de gangbare verklaring, dat myelinisatie een middel is om de geleidingssnelheid over lange afstanden te verhogen, niet onmiddellijk van toepassing. Lokale collateralen behoeven geen (?) vergroting ten opzichte van veraf gelegen doelen en in feite, voor de synchroniciteit die men had kunnen voorspellen, lijkt een versterkte lokale geleiding (via myelinisatie?) paradoxaal. Zou het kunnen dat de myelinisatie verband houdt met andere factoren, zoals plasticiteits-gerelateerde (of toestand-gerelateerde) veranderingen in axondiameter?
Een interessant voorbeeld: Meynert Cellen in Gebied V1 van niet-menselijke Primaten
Deze grote neuronen, op de grens van lagen 5 en 6, projecteren variabel naar extrastriate gebied MT, en/of andere visuele associatie gebieden, en/of naar de pulvinar kern, en/of naar de superieure colliculus en pretectum (Weisenhorn et al., 1995; Rockland en Knutson, 2001). De intrinsieke collateralisatie is uitzonderlijk uitgebreid, gemeten als 8,0 mm van het soma, op basis van subtotale reconstructies, en met ten minste 800-1.370 boutons (figuur 1). Over de axonale arborisatie, lijkt er een duidelijke variabiliteit in tak diameter, dat wil zeggen, extrinsieke takken gericht op gebied MT zijn groot (diameter ~ 3,0 urn), maar de intrinsieke takken en die projecteren naar de pulvinar en superieure colliculus lijken kleiner, zoals beoordeeld door lichtmicroscopie (cf. Figuur 1B (intrinsieke) en Figuur 1D (extrinsieke naar gebied MT). Verschillende diameters geven vermoedelijk verschillen in de mate van myelinisatie en, bij afleiding, in geleidingssnelheid.
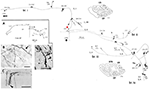
Figuur 1. Een typische, ruimtelijk uitgebreide proximale axonale arborisatie van een Meynert-cel (rode asterisk) in de primaire visuele cortex van een makaakaap. Er zijn drie grote intrinsieke collateralen (aangeduid als br. i, ii, iv), die 3,0 mm dorsaal in laag 1, 3,0 mm ventraal in laag 6, 4,0 mm ventraal in laag 6, en 0,5 mm in laag 4B, zoals aangegeven door pijlpunten in de twee coronale doorsnede contouren (secties 222 en 274, waar dorsaal is naar links). Alles bij elkaar beslaan de intrinsieke collateralen 5,9 mm anterior-posterior (117 coupes × 50 μm). Een andere, extrinsieke collateraal (br. iii) komt voor in lagen 4 en 6 van gebied V2. Gedeelten van de afzonderlijke collateralen en van het hoofdaxon (dikke pijl) konden niet worden gevolgd, zoals is aangegeven met stippellijnen. De lage vergrotingsinset (A, links) geeft een schematisch overzicht van de algemene configuratie. Nummers geven individuele secties, waarbij 20 nummers = 1,0 mm. Alle takken hebben talrijke kleine synaptische clusters, waarvan er een wordt geïllustreerd in (B). Let op de verminderde diameter tussen de belangrijkste axon (pijl) en de terminal arborization. Extrinsieke axonen (C) zijn van variabele diameter (een grote diameter axon bij de pijlpunt). (D) Extrinsieke beëindigingen in gebied MT omvatten enkele grote diameter axonen. Schaalstreep = 25 μm in (B), 100 μm in (C), 20 μm in (D). CF, calcarine fissure; LS, lunate sulcus; STS, superior temporal sulcus; L, laag. Gewijzigd van figuren 1, 9 in Rockland en Knutson (2001) en figuren 6f, 7b van Rockland (1995) met toestemming.
Functionele betekenis van axon vertakking
Een belangrijke component van collateralisatie is dat de dochter takken zijn vaak niet uniform, maar vooral op vertakkingspunten, variëren in diameter (figuur 2). Variabiliteit in diameter samen met andere parameters zal invloed hebben op prikkelbaarheid, geleidingssnelheid, en andere aspecten van de signaalvoortplanting. Andere parameters die van invloed zijn, zijn onder meer de breedte van myeline en intermodale lengte, en de dichtheid en distributie van ionenkanalen (besproken in Debanne et al., 2011; Seidl, 2014; Bucher, 2016; Seidl en Rubel, 2016; Rama et al., 2018). Deze zouden effecten hebben op neurale responseigenschappen op microcircuitniveau. Op het meer globale niveau zou gesommeerde activiteit van meerdere projecterende axonen, met variërende geleidingssnelheden over een onderling verbonden netwerk, kunnen resulteren in een spectrum van synchrone en/of asynchrone activaties (Mitra et al., 2015; Zeki, 2016). Differentiële rekrutering van postsynaptische populaties of netwerkrecombinaties zouden factoren kunnen zijn in toestandsovergangen of modulatie.
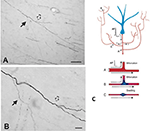
Figuur 2. Een typisch vertakt segment van een axon in de witte stof (makaakaap). Het segment is afkomstig van een neuron in de pariëtale cortex en is hier te zien in de nabijheid van de ventrale temporale cortex. Paneel (A) is een lagere vergroting van (B). Let op de dubbele bifurcaties, waarbij de eerste dochter tak (vaste pijl) opvallend dunner (en ongemyeliniseerd?) is dan het hoofd axon. In de tweede, iets meer distale bifurcatie (holle pijl), lijken de dochtervertakkingen ongeveer even groot in diameter, maar beide zijn dunner dan het hoofdaxon. Schaalstaven = 100 micrometer (A) en 10 micrometer (B). Overgenomen van Zhong en Rockland (2003) met toestemming. (C) Schematische weergave van een neuron (blauw) en de uitgebreide vertakking topologie (verkort omwille van handige opmaak). Een actiepotentiaal (AP) kan circuitvormige paden (A-C) volgen naar meerdere doelen. Betrouwbaarheid van de voortplanting hangt af van zowel de actieve elektrische eigenschappen van het axon en de geometrie, met inbegrip van membraan inhomogeneties zoals zwellingen en onverenigbare tak diameters. Hieronder: schema ter illustratie van betrouwbare voortplanting (A, met optimale impedantieovereenstemming tussen de moeder- en de dochtertak), en vertraagde of mislukte voortplanting (B, wanneer de dochtertak een vergrote diameter heeft; C, wanneer er een tussenliggende membraanzwelling is). Overgenomen uit Huguenard (2000) met toestemming.
De axongeometrie, actieve elektrische eigenschappen, en membraan inhomogeniteiten op vertakkingspunten zijn bekend als factoren van betrouwbare propagatie (bijv. Manor et al., 1991; Innocenti et al., 1994; Tettoni et al., 1998; Huguenard, 2000; Ofer et al., 2017). Dit leidt tot verschillende scenario’s over temporele kenmerken die het gevolg zijn van collateralisatie.
Ten eerste kan er sprake zijn van synchrone activatie door de dochtervertakkingen. De auditieve hersenstam pathways projecteren via een enkel bifurcerende axon naar ipsi- en contralaterale doelen (respectievelijk korte en langere fysieke pathways). Isochronische transmissie wordt bereikt door differentiële myelinisatie en axonkaliber van de twee dochtertakken (d.w.z. kleiner kaliber en kortere intermodale lengtes ipsilateraal; Seidl, 2014; Seidl en Rubel, 2016). Aangezien de meeste vertakte axonen, in vergelijking met de hersenstam auditieve paden, een groter gebied bestrijken en minder goed gedefinieerde functies dienen, zijn gegevens grotendeels onvolledig of ontbreken voor andere systemen. Men zou echter kunnen voorspellen, in het geval van synchroniciteit, dat proximale takken (d.w.z. de intrinsieke collateralen van Meynert of andere piramidale cellen) anatomische specialisaties zouden hebben die resulteren in langere geleidingstijden, om te compenseren voor en te matchen met de langere afstanden van extrinsieke collateralen. Zoals hierboven opgemerkt, lijkt deze eenvoudige voorspelling niet op te gaan. Verder onderzoek zal bemonstering van geïdentificeerde axonen over lange afstanden met zich meebrengen, en zou niet gemakkelijk te realiseren zijn.
Tweede, tak-specifieke activering kan asynchroon zijn. Dit kan te wijten zijn aan selectieve uitval van transmissie en / of asynchrone geleidingstijden over de axon arbor (figuur 2; Huguenard, 2000; Bucher, 2016). Modellen van corticale circuits beschrijven onderscheidende routeringstoestanden van kortstondige voorbijgaande synchronie die dynamisch vorm zouden kunnen geven aan de informatiestroom (Palmigiano et al., 2017). Vergelijkbare experimentele gegevens ontbreken grotendeels voor lange afstand axonen. De collaterale topologie van thalamocorticale projecties biedt echter één voorbeeld dat evocatief is voor een gecompliceerd, activiteits- en/of toestandsafhankelijk asynchroon activeringspatroon.
Corticale en thalamocorticale activiteit is sterk toestandsafhankelijk; en de interactie van presynaptische extrinsieke inputs (tak specifiek?) met intrinsieke membraan- en synaptische eigenschappen van postsynaptische neuronen wordt beschouwd als fundamenteel voor het genereren van ritmische activiteit (met “breed scala aan effecten van het versterken of blokkeren van sensomotorische verwerking….,” McCormick et al., 2015).
Variabiliteit in corticale reacties is paradoxaal omdat deze ook dienen als substraten van stabiele zintuiglijke ervaring. Neuronale variabiliteit is geassocieerd met de mate van heterogene synchronie over extrinsieke input; dat wil zeggen, zintuiglijke stimulatie of gedragscondities die de homogeniteit van de input naar een bepaald gebied verhogen, wordt voorspeld dat ze ook de neuronale variabiliteit verminderen (Gomez-Laberge et al., 2016). Recente discussies over microcircuitreacties hebben gespeculeerd over een prominente rol van lichte varianties of verschil in informatie: “Maar wat als de verschillen tussen de connectiviteit binnen cohorten van cellen van dezelfde klasse belangrijk zijn voor de circuitfunctie?” (Morgan en Lichtman, 2017).
Dynamische Axon Eigenschappen
Verschuivingen in responslatentie zijn gerapporteerd in relatie tot verschillende staten van alertheid. In de corticothalamus pathway resulteert verhoogde alertheid in een significant kortere reactietijd. Dit en/of veranderingen in de afvuurfrequentie van aankomende impulsen kunnen verantwoordelijk zijn voor een dramatisch verhoogde responsbetrouwbaarheid voor de subpopulatie (58%) van visueel responsieve corticogeniculate neuronen (in konijnen: Stoelzel et al., 2017). Deze resultaten hebben betrekking op fysiologisch geïdentificeerde enkele axonen; maar men kan speculeren over een bredere toepasbaarheid op takken van collateralized axonen.
Ongrijpende processen van synaptogenese en distale axon turnover zijn aangetoond in de volwassen cortex (NHP: Stettler et al., 2006). Op kortere tijdschalen toont superresolutiemicroscopie van ongemyeliniseerde GFP-gelabelde CA3 hippocampus in organotypische hersenschijfjes aan dat axonen geleidelijk verbreden na vlagen van hoogfrequente vuren, een observatie bevestigd door elektrofysiologische opname (Chereau et al., 2017). Andere, tak-specifieke veranderingen zijn waarschijnlijk te ontdekken; bijvoorbeeld, terminal arborizations van individueel gelabelde axonen uit de dorsale raphe hebben een doel-specifiek percentage van boutons die het eiwit VGLUT3 bevatten (groter percentage voor takken die eindigen in het striatum dan in de motorische cortex). Dit impliceert een complex, niet-uniform trafficking mechanisme over collateralen (Gagnon en Parent, 2014).
Conclusie
In dit Perspectief artikel, heb ik axonvertakking besproken als relevant voor veranderingen in hersentoestand, met impact via tak-specifieke eigenschappen, differentiële rekrutering van postsynaptische ensembles, en hele hersenen patronen van synchronisatie. Dit bouwt voort op lange-termijn discussies over axonale vertakking topologieën en hoe deze informatieverwerking kunnen moduleren door tijdsvertragingen in impulsvoortplanting, differentiële tak-specifieke filtering, en activiteit-afhankelijke prikkelbaarheid (e.g., Segev en Schneidman, 1999). Met slechts enkele uitzonderingen, zoals de auditieve hersenstam pathway, ontbreken harde gegevens nog grotendeels over synchrone en asynchrone activaties door dochtertakken en hoe deze temporele relaties van invloed zouden kunnen zijn op postsynaptische neuronale responsiviteit (maar, zie Gomez-Laberge et al., 2016; Stoelzel et al., 2017). Een voortdurende uitdaging is dus het ophelderen van tak-specifieke kenmerken binnen individuele axonen en de effecten op postsynaptische ensembles. Recent werk brengt aanvullende vragen over netwerk heterogeniteit naar voren, waaronder waarom neuronen uit een enkel brongebied variabel projecteren naar een of meer doelen in wat herhaaldelijk wordt beschreven als “in alle combinaties.”
Author Contributions
De auteur bevestigt de enige bijdrager van dit werk te zijn en het goed te keuren voor publicatie.
Funding
We danken National Institutes of Health (NIH) voor financieringssteun: MH107456.
Conflict of Interest Statement
De auteur verklaart dat het onderzoek werd uitgevoerd in de afwezigheid van commerciële of financiële relaties die kunnen worden opgevat als een potentieel belangenconflict.
Barry, J. M. (2015). Axonale activiteit in vivo: technische overwegingen en implicaties voor de exploratie van neurale circuits in vrij bewegende dieren. Front. Neurosci. 9:153 doi: 10.3389/fnins.2015.00153
PubMed Abstract | CrossRef Full Text | Google Scholar
Gagnon, D., and Parent, M. (2014). Distribution og VGLUT3 in highly collateralized axons from the rat dorsal raphe nucleus as revealed by single-neuron reconstructions. PLoS ONE 9:e87709. doi: 10.1371/journal.pone.0087709
CrossRef Full Text | Google Scholar
Kita, T., and Kita, H. (2012). The subthalamic nucleus is one of multiple innervation sites for long-range corticofugal axons: a single axon tracing study in the rat. J. Neurosci. 32, 5990-5999 doi: 10.1523/JNEUROSCI.5717-11
PubMed Abstract | CrossRef Full Text | Google Scholar