Frontiers in Systems Neuroscience
Az agyi állapot változásai a neuronok tüzelési tulajdonságainak mikroáramköri változásaihoz és az agyi aktiváció szinkron vagy aszinkron mintázatának makroszintű változásaihoz kapcsolódnak. Ebben a keretben az axonokat viszonylag kevésbé vizsgálták (Barry, 2015), és akkor is elsősorban az akciós potenciálok vezetési sebességével és a vezetési sebesség változásaiból eredő fokozott éberséggel összefüggésben (pl. Stoelzel et al., 2017). Ezen túlmenően azonban sok axon erősen elágazó topológiája jól megtervezettnek tűnik ahhoz, hogy differenciáltan rekrutálja az elosztott posztszinaptikus neuroncsoportokat, valószínűleg állapotfüggő módon. A jelfeldolgozás legalább három axonális tartományban történhet; nevezetesen a leképezés, az erősítés és az időzítés (Innocenti et al., 2016; Innocenti, 2017). Ebben a perspektívában először röviden megvitatom az axon-kollateralizáció hálózati példáit, majd néhány axon-intrinsic jellemzőt, amelyek a differenciált posztszinaptikus rekrutáció hátterében állhatnak. Részletes paraméterek vagy mechanizmusok hiányában a cél elsősorban az általános jellemzők kiemelése, amelyek az agyi állapotok szabályozásában és átmeneteiben szerepelhetnek.
Axon kollaterálisok
Minden axonnak van egy kidolgozott disztális arborizációja a célstruktúrában. Sok axonnak ráadásul több elágazása (más néven kollaterálisa) van, amelyek markánsan különböző struktúrákat céloznak meg (áttekintve: Rockland, 2013, 2018). Kiváló példa erre az 5. rétegbeli kortikotalamikus neuronok axonjainak elágazása. Ezt többször dokumentálták már, és gyakran tárgyalják, mint olyan kulcsfontosságú mechanizmust, amelynek segítségével a szervezet meg tudja különböztetni, hogy az érzékszervi bemenet változásait a környezet változása vagy a saját maga által kezdeményezett mozgások okozzák-e (“efferenciamásolat” vagy “kollaterális kisülés”, Guillery és Sherman, 2011).
Rágcsálókban, ahol számos alkalmazható technika áll rendelkezésre e kérdés vizsgálatára, a kollateralizáció viszonylag gyakori. Hogy néhány példát említsünk: (1) A hippokampusz subiculum neuronjai a mammilláris testekbe, a retrosplenialis kéregbe vagy kollaterálisok révén mindkettőbe vetülnek (patkány: Kinnavane et al, 2018); (2) a dorsalis raphe (DR) magban lévő szerotonerg neuronok különböző kombinációkban érintkeznek a striatummal, a prefrontális kéreggel és az amygdalával (patkány: Gagnon és Parent, 2014), valamint a vegetatív magok kombinációival (Waselus et al., 2011). Ebben az esetben a stresszválasz DR-kollaterálisok általi aktiválása elérheti a neurohormonok felszabadulásával vagy a presszorválaszokkal kapcsolatos magok szinkronizált aktiválását. Az előagyi struktúrák differenciált, koordinált aktivációi hozzájárulhatnak a DR sokrétű, de egymással összefüggő funkcióihoz, például az alvás-ébrenlét ciklus szabályozásához, a fájdalomjelek modulációjához vagy a hangulat kifejeződéséhez (Gagnon és Parent, 2014).
Harmadszor, az egér thalamocorticalis projekciók anatómiai vizsgálatai “multispecifikus axonokat” azonosítanak, amelyek széles körben ágaznak el a különálló agykérgi (és szubkortikális) területek korlátozott tartományaiba. Feltételezték, hogy ezek hangszerelik a térben elosztott, szinkronizálható neurális összeállítások gyors kialakulását és újrakonfigurálását (Clasca et al., 2016).
Negyedszer, a kortikokortikális kapcsolatok egész agyi axonális nyomkövetéssel végzett közelmúltbeli vizsgálata egér vizuális kéregben azt találta, hogy 30 neuronból 23 két-hét másik kortikális területről lépett kapcsolatba. Ennek az eredménynek a megerősítéseként a genetikailag vonalkódolt neuronok nagy áteresztőképességű DNS-szekvenálása 533 neuron 44%-át találta többszörösen kivetülőnek (“broadcast neurons”, Han et al., 2018). Han és munkatársai ideiglenesen a kivetülő neuronok két széles típusát különböztették meg, egy kisebb “dedikált” (egycélú) alpopulációt, amely a “sugárzó” (többszörösen kivetülő) sejtek túlsúlyával együtt létezik. Lehet, hogy ez az architektúra a kognitív állapot és az érzékszervi feldolgozás modulációit szolgálja?
Nem sztereotipizált kollateralizáció
Amint fentebb megjegyeztük, a kollaterálódó neuronok nem sztereotipizált mintázatban teszik ezt. Egy kijelölt (az eredet által meghatározott) projekción belül a neuronok a célterületek változatos részhalmazába (“minden kombinációban”) küldenek elágazásokat. Ez a megfigyelés annyira konzisztens, hogy szabályként tekinthetjük, bár a részletes paramétereket nem táblázatosítottuk. A funkcionális jelentősége szintén nem világos, de a felmerülő eredmények azt sugallják, hogy a heterogén és változó bemenetek egy agykérgi neuronhoz (és, kiterjesztve, következtethetünk az idegi összeszerelésekre) fontosak a variabilitás és a spike train változásainak vezetésében a kísérleti próbák során (Gomez-Laberge et al, 2016 és lásd alább).
Intrinsic Collaterals
A hosszú távú agykérgi projekciós neuronok minden fajban, az egyetlen vagy többszörös extrinsic célpontok mellett, jellemzően kidolgozott intrinsic arborizációval rendelkeznek. Egyes agykérgi neuronok csak intrinsic (lokális) kollaterizációval rendelkeznek, mások pedig csak extrinsic (patkány: Kita és Kita, 2012). Bár az intrinsic és extrinsic arborizáció tényleges arányaira vonatkozóan csak kevés adat áll rendelkezésre (Parent et al., 2000; Rockland, 2018), a bizonyítékok arra utalnak, hogy ez nagyon változó lesz. Még az intrinsic kapcsolatok rendszerén belül is nemcsak a kollaterálisok és a szinaptikus boutonok száma között vannak különbségek, hanem egyetlen neuron is rendelkezhet myelinizált és nem myelinizált kollaterálisok keverékével (macska látókéreg: Martin et al., 2014; Koestinger et al., 2017). Az ágspecifikus mielinizáció megnövekedett, ágspecifikus vezetési sebességet eredményezhet, bár Koestinger és munkatársai szerint ennek inkább olyan tényezőkhöz lehet köze, mint a fokozott átviteli biztonság, feltehetően ismét ágspecifikus.
A mielinizációval kapcsolatos különös megfigyelés a Gennari stria-ra, a főemlős V1 terület 4B rétegében lévő axonok mielinizált sávjára vonatkozik. Mivel ez intrinsic kollaterálisokból áll, az általános magyarázat, miszerint a myelinizáció a vezetési sebesség növelésének eszköze nagy távolságokon keresztül, nem alkalmazható azonnal. A helyi kollaterálisoknak nincs szükségük (?) növelésre a távoli célpontokhoz képest, és valójában a szinkronitás, amelyet előre lehetett volna jelezni, a fokozott helyi vezetés (a myelinizáción keresztül?) paradoxnak tűnik. Lehet, hogy a myelinizáció más tényezőkhöz kapcsolódik, például az axon átmérőjének plaszticitással kapcsolatos (vagy az állapottal kapcsolatos) változásaihoz?
Egy érdekes példa: Meynert-sejtek a főemlősök V1-es területén
Ezek a nagyméretű neuronok az 5. és 6. réteg határán variálhatóan az extrastriatális MT területre és/vagy más vizuális asszociációs területekre és/vagy a pulvináris magra és/vagy a colliculus superiorra és a pretectumra vetülnek (Weisenhorn és mtsai., 1995; Rockland és Knutson, 2001). Az intrinsic kollateralizáció kivételesen kiterjedt, a szubtotális rekonstrukciók alapján 8,0 mm-re mérve a soma-tól, és legalább 800-1 370 boutonnal rendelkezik (1. ábra). Az axonális arborizáció mentén az ágak átmérőjében határozott variabilitás mutatkozik; azaz az MT területre irányuló extrinsic ágak nagyok (átmérő ~3,0 μm), de az intrinsic ágak és a pulvinar és a superior colliculus felé kivetülő ágak kisebbnek tűnnek, fénymikroszkópos vizsgálat alapján (vö. 1B ábra (intrinsic) és 1D ábra (extrinsic az MT területre). A változó átmérők feltehetően a myelinizáció mértékében és ebből következően a vezetési sebességben mutatkozó különbségekre utalnak.
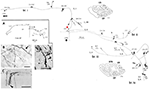
1. ábra. Egy Meynert-sejt (piros csillag) tipikus, térben kiterjedt proximális axonális arborizációja makákó majom elsődleges látókéregében. Három fő intrinzikális kollaterális (br. i, ii, iv jelöléssel) van, amelyek az 1. rétegben dorsalisan 3,0 mm-re, a 6. rétegben ventrálisan 3,0 mm-re, a 6. rétegben ventrálisan 4,0 mm-re, a 4B rétegben pedig 0,5 mm-re nyúlnak, amint azt a két koronális metszet vázlatában (222. és 274. metszet, ahol a dorsális balra van) nyílhegyek jelzik. A belső kollaterálisok együttesen 5,9 mm-t tesznek ki elöl-hátul (117 metszet × 50 μm). Egy további, extrinsic kollaterális (br. iii) a V2 terület 4. és 6. rétegében található. Az egyes kollaterálisok és a fő axon (vastag nyíl) egyes részeit nem lehetett követni, amit szaggatott vonalak jeleznek. Az alacsony nagyítású betét (A, balra) sematikus áttekintést nyújt az általános konfigurációról. A számok az egyes metszeteket jelölik, ahol 20 szám = 1,0 mm. Minden ágon számos kis szinaptikus klaszter található, amelyek közül az egyiket a (B) ábrázolja. Figyeljük meg a fő axon (nyíl) és a terminális arborizáció közötti csökkent átmérőt. A külső axonok (C) változó átmérőjűek (egy nagy átmérőjű axon a nyílhegynél). (D) Az MT területen lévő extrinsic terminációk néhány nagy átmérőjű axont tartalmaznak. Méretsáv = 25 μm (B), 100 μm (C), 20μm (D). CF, calcarine fissure; LS, lunate sulcus; STS, superior temporal sulcus; L, réteg. Rockland és Knutson (2001) 1., 9. ábrája és Rockland (1995) 6f, 7b ábrája alapján, engedéllyel módosítva.
A tengelyelágazás funkcionális jelentősége
A kollateralizáció fontos összetevője, hogy a leányágak gyakran nem egységesek, hanem különösen az elágazási pontokon eltérő átmérőjűek (2. ábra). Az átmérő változékonysága más paraméterekkel együtt hatással van az ingerelhetőségre, a vezetési sebességre és a jelterjedés más aspektusaira. További befolyásoló paraméterek közé tartozik a mielin szélessége és az intermodális hossz, valamint az ioncsatornák sűrűsége és eloszlása (áttekintve: Debanne et al., 2011; Seidl, 2014; Bucher, 2016; Seidl és Rubel, 2016; Rama et al., 2018). Ezek hatással lennének az idegi válasz tulajdonságaira a mikrokörök szintjén. Globálisabb szinten több kivetülő axon összegzett aktivitása, különböző vezetési sebességekkel egy összekapcsolt hálózaton keresztül, a szinkron és/vagy aszinkron aktivációk spektrumát eredményezheti (Mitra et al., 2015; Zeki, 2016). A posztszinaptikus populációk differenciált toborzása vagy hálózati rekombinációk lehetnek az állapotátmenetek vagy a moduláció tényezői.
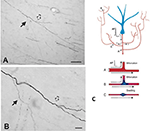
2. ábra. Egy tipikusan elágazó axonszakasz a fehér anyagban (makákó majom). A szegmens a parietális kéreg egyik neuronjából ered, és itt a ventrális temporális kéreg közelében látható. Az (A) panel a (B) kisebb nagyítása. Figyeljük meg a kettős elágazásokat, ahol az első leányág (tömör nyíl) feltűnően vékonyabb (és nem myelinizált?), mint a fő axon. A második, kissé távolabbi elágazásban (üreges nyíl) a leányágak átmérője körülbelül azonosnak tűnik, de mindkettő vékonyabb, mint a fő axon. Méretsávok = 100 μm (A) és 10 μm (B). Zhong és Rockland (2003) engedélyével másolva. (C) Egy neuron (kék) és annak kiterjesztett elágazási topológiájának sematikus ábrája (a kényelmes formázás érdekében előrerövidítve). Egy akciós potenciál (AP) kerülő utakat (A-C) követhet több célponthoz. A terjedés megbízhatósága az axon aktív elektromos tulajdonságaitól és geometriájától is függ, beleértve a membrán inhomogenitásait, mint például a duzzanatok és az inkompatibilis ágátmérők. Az alábbiakban: a megbízható terjedést (A, optimális impedanciaillesztés az anya és a leányágak között), valamint a lelassult vagy sikertelen terjedést (B, ahol a leányág átmérője megnövekedett; C, ahol a membrán duzzanata van közöttük) szemléltető ábra. Huguenard (2000) engedélyével reprodukálva.
Az axon geometriája, az aktív elektromos tulajdonságok és a membrán inhomogenitásai az elágazási pontokon jól ismert tényezői a megbízható terjedésnek (pl. Manor et al., 1991; Innocenti et al., 1994; Tettoni et al., 1998; Huguenard, 2000; Ofer et al., 2017). Ez több különböző forgatókönyvhöz vezet a kollateralizációból következő időbeli jellemzőkkel kapcsolatban.
Először, szinkron aktiváció lehet az egész leányágban. A halló agytörzsi pályák egyetlen elágazó axonon keresztül vetülnek ipsi- és kontralaterális célpontokhoz (illetve rövid és hosszabb fizikai pályákhoz). Az izokron átvitel a két leányág eltérő myelinizációjával és axonkaliberével érhető el (azaz kisebb kaliberű és rövidebb intermodális hosszúságú ipsilaterálisan; Seidl, 2014; Seidl és Rubel, 2016). Mivel a legtöbb elágazó axon az agytörzsi hallójáratokhoz képest nagyobb területet fed le és kevésbé jól meghatározott funkciókat szolgál, az adatok nagyrészt hiányosak vagy hiányoznak más rendszerekre vonatkozóan. A szinkronitás esetében azonban előre jelezhetnénk, hogy a proximális ágak (azaz a Meynert vagy más piramissejtek intrinzikális kollaterálisai) anatómiai specializációval rendelkeznek, ami hosszabb vezetési időt eredményez, hogy kompenzálják az extrinsic kollaterálisok hosszabb távolságait és illeszkedjenek azokhoz. Mint fentebb említettük, úgy tűnik, ez az egyszerű előrejelzés nem állja meg a helyét. A további vizsgálatok az azonosított axonokból való mintavételt igénylik hosszú távolságokon keresztül, és nem lenne könnyű megvalósítani.
Második, az ágspecifikus aktiváció aszinkron lehet. Ennek oka lehet az átvitel szelektív meghibásodása és/vagy aszinkron vezetési idők az axon arboron keresztül (2. ábra; Huguenard, 2000; Bucher, 2016). Az agykérgi áramkörök modelljei leírják a rövid életű átmeneti szinkronitás megkülönböztető útirányítási állapotait, amelyek dinamikusan alakíthatják az információáramlást (Palmigiano et al., 2017). Összehasonlítható kísérleti adatok nagyrészt hiányoznak a hosszú távú axonok esetében. A thalamocorticalis projekciók kollaterális topológiája azonban egy olyan példát nyújt, amely egy bonyolult, aktivitás- és/vagy állapotfüggő aszinkron aktivációs mintázatot idéz.
A kortikális és thalamocorticalis aktivitás erősen állapotfüggő; és a preszinaptikus extrinsic bemenetek kölcsönhatása (ágspecifikus?) a posztszinaptikus neuronok intrinsic membrán és szinaptikus tulajdonságaival alapvetőnek tekinthető a ritmikus aktivitás generálásában (a “széleskörű hatások a szenzomotoros feldolgozás fokozásától vagy blokkolásától kezdve…., McCormick et al., 2015).
A kortikális válaszok változékonysága paradoxon, mivel ezek a stabil érzékszervi tapasztalatok szubsztrátjaiként is szolgálnak. A neuronális variabilitást összefüggésbe hozták a heterogén szinkronitás mértékével az extrinsic inputon; vagyis az olyan szenzoros stimuláció vagy viselkedési körülmények, amelyek növelik az adott területre érkező input homogenitását, az előrejelzések szerint a neuronális variabilitást is csökkentik (Gomez-Laberge et al., 2016). A mikroáramkörök válaszainak közelmúltbeli vitái a kis eltérések vagy az információk közötti különbség kiemelkedő szerepéről spekuláltak: “De mi van akkor, ha az azonos osztályba tartozó sejtek kohorszain belüli konnektivitás közötti különbségek fontosak az áramkör működése szempontjából?”. (Morgan és Lichtman, 2017).
Dinamikus axontulajdonságok
A válaszlatencia változásairól számoltak be a különböző éberségi állapotokkal kapcsolatban. A kortikotalamikus pályán a fokozott éberség jelentősen lerövidült válaszadási késleltetést eredményez. Ez és/vagy a beérkező impulzusok tüzelési frekvenciájának változása lehet felelős a vizuálisan reagáló kortikogenuláris neuronok alpopulációjának (58%) drámaian megnövekedett válaszmegbízhatóságáért (nyulakban: Stoelzel et al., 2017). Ezek az eredmények fiziológiailag azonosított egyes axonokra vonatkoznak; de feltételezhető egy szélesebb körű alkalmazhatóság a kollaterális axonok ágaira.
A szinaptogenezis és a disztális axonforgalom folyamatos folyamatait kimutatták a felnőtt kéregben (NHP: Stettler et al., 2006). Rövidebb időskálán, organotípusos agyszeletekben a nem myelinizált GFP-vel jelölt CA3 hippokampuszok szuperfelbontású mikroszkópiája azt mutatja, hogy az axonok fokozatosan kiszélesednek a nagyfrekvenciás tüzelési rohamok után, ezt a megfigyelést elektrofiziológiai felvételek is megerősítették (Chereau et al., 2017). Valószínűleg más, ágspecifikus változások is felfedezhetők; például a dorsalis raphe-ból származó, egyedileg jelölt axonok terminális arborizációi a VGLUT3 fehérjét tartalmazó boutonok célspecifikus százalékos arányával rendelkeznek (nagyobb százalék a striatumban végződő ágak esetében, mint a motoros kéregben). Ez egy összetett, nem egyenletes forgalmi mechanizmust feltételez a kollaterálisokon keresztül (Gagnon és Parent, 2014).
Következtetés
Ebben a perspektivikus cikkben az axonok elágazását az agyi állapot változásai szempontjából relevánsnak tárgyaltam, amelynek hatása az ágspecifikus tulajdonságokon, a posztszinaptikus együttesek differenciált rekrutációján és az egész agyi szinkronizációs mintázatokon keresztül érvényesül. Ez az axonális elágazási topológiákkal kapcsolatos hosszú távú vitákra épül, és arra, hogy ezek hogyan modulálhatják az információfeldolgozást az impulzusok terjedésének időbeli késleltetése, a differenciális ágspecifikus szűrés és az aktivitásfüggő gerjeszthetőség révén (pl. Segev és Schneidman, 1999). Néhány kivételtől eltekintve, mint például a halló agytörzsi pálya, a kemény adatok még mindig nagyrészt hiányoznak a leányágakon keresztül történő szinkron és aszinkron aktivációkról, és arról, hogy ezek az időbeli kapcsolatok hogyan befolyásolhatják a posztszinaptikus neuronális válaszkészséget (de lásd Gomez-Laberge et al., 2016; Stoelzel et al., 2017). Így folyamatos kihívást jelent az egyes axonokon belüli ágspecifikus jellemzők és a posztszinaptikus együttesekre gyakorolt hatások tisztázása. A legújabb munkák további kérdéseket vetnek fel a hálózati heterogenitással kapcsolatban, többek között azt, hogy az egyetlen forrásterületről származó neuronok miért vetülnek változóan egy vagy több célpontra, amit ismételten úgy írnak le, hogy “minden kombinációban.”
A szerző hozzájárulása
A szerző megerősíti, hogy ő a munka egyedüli szerzője, és jóváhagyta azt a közzétételre.
Finanszírozás
Köszönjük a National Institutes of Health (NIH) finanszírozási támogatását: MH107456.
Conflict of Interest Statement
A szerző kijelenti, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végeztük, amelyek potenciális összeférhetetlenségként értelmezhetők.
Barry, J. M. (2015). Axonális aktivitás in vivo: technikai megfontolások és következmények az idegi áramkörök feltárására szabadon mozgó állatokban. Front. Neurosci. 9:153 doi: 10.3389/fnins.2015.00153
PubMed Abstract | CrossRef Full Text | Google Scholar
Gagnon, D., and Parent, M. (2014). A VGLUT3 og eloszlása a patkány háti raphe magból származó, erősen kollaterális axonokban, amint azt az egyneuronos rekonstrukciók feltárják. PLoS ONE 9:e87709. doi: 10.1371/journal.pone.0087709
CrossRef Full Text | Google Scholar
Kita, T., and Kita, H. (2012). A subthalamicus mag egyike a hosszú távú kortikofugális axonok többféle innervációs helyének: Egyetlen axon nyomkövetési vizsgálata patkányban. J. Neurosci. 32, 5990-5999 doi: 10.1523/JNEUROSCI.5717-11
PubMed Abstract | CrossRef Full Text | Google Scholar